
Entry Version:
Citation:
Pancreapedia: Exocrine Pancreas Knowledge Base, DOI: 10.3998/panc.2020.14
| Attachment | Size |
|---|---|
| 709.75 KB | |
| 1.05 MB |
I. INTRODUCTION
The exocrine pancreas secretes digestive enzymes, fluid, and bicarbonate in response to food ingestion. This is a critical digestive process that is regulated by neural reflexes, gastrointestinal hormones, and absorbed nutrients. Secretion is highly regulated by both stimulatory and inhibitory influences that coordinate the delivery of digestive enzymes with food emptying into the intestine to assure adequate digestion of a meal. In the absence of proper pancreatic secretion, maldigestion and malabsorption of nutrients may cause malnutrition and associated complications. This review will describe the physiological processes that regulate pancreatic exocrine secretion.
II. PHASES OF MEAL RESPONSE
Pancreatic secretion in response to a meal occurs in four distinct but overlapping phases which are named based on the location of ingested food. The four phases of pancreatic secretion are cephalic, gastric, intestinal, and absorbed nutrient. Considerable crosstalk and inter-regulation is associated within the phases, thereby ensuring adequate, but not excessive, enzyme and bicarbonate secretion. Each phase is regulated by a complex network of neural, humoral, and paracrine feedback mechanisms which help to maintain an optimal environment for food digestion and absorption.
A. Cephalic Phase
Sensory inputs such as sight, smell, taste, and mastication (prior to swallowing) lead to the anticipation of food. These sensations initiate the first phase of pancreatic secretion known as the cephalic phase. In addition to sensory input, interaction of certain food molecules such as long chain fatty acids (but not triglycerides or medium chain fatty acids) with receptors in the oral cavity also induce the cephalic phase (125). Furthermore, studies in animals have implicated a gustatory vago-pancreatic reflex in mediating the cephalic phase of pancreatic secretion (259, 284). Approximately 20-25% of the total pancreatic exocrine secretion occurs during the cephalic phase (9, 59, 173). This estimate is based on data obtained by sham feeding, a process by which food is anticipated by sight, smell, and taste, but not ingested. Sham feeding in animals such as dogs, has been evaluated by inserting a surgically prepared gastric fistula that diverts food from the esophagus, allowing swallowing but not entry of food into the stomach. In humans, sham feeding involves chewing but not swallowing. The pancreatic response to sham feeding in humans lasts approximately 60 minutes while in dogs it can last for more than 4 hours (301, 316). Sham feeding stimulates pancreatic secretion which is low in bicarbonate but rich in enzymes, suggesting that pancreatic acinar, rather than ductal cells are stimulated in this phase (9).
The cephalic phase of exocrine secretion is under the control of the vagus nerve. Sensory inputs arising from anticipation of food are integrated in the dorsal vagal complex (located in the brainstem) and transmitted to the exocrine pancreas via the vagus nerve (88, 269). Cholinergic agonists produce secretory responses similar to cephalic stimulation while vagotomy blocks the cephalic responses, suggesting that acetylcholine released by vagal efferents is the primary mechanism by which sensory inputs lead to exocrine secretion (20, 131). Secretion of the islet hormone, pancreatic polypeptide (PP), increases with sham feeding and serves as an indicator of vagal innervation of the pancreas, as its secretion is inhibited by cholinergic blockers (159, 307). When sham feeding is accompanied by swallowing, the pancreatic secretory and PP responses are much greater implying that chewing and swallowing stimulate PP secretion by cholinergic mechanisms (308).
The exocrine pancreas also contains peptidergic nerve terminals and there is some evidence to suggest that neuropeptides such as vasoactive intestinal peptide (VIP) and gastrin-releasing peptide (GRP) may influence the cephalic phase. In addition, thyrotropin-releasing hormone stimulates pancreatic exocrine secretion of protein and bicarbonate through vagal efferents and this process involves both muscarinic and VIP receptors (9, 127, 170). In contrast, the effects of inhibitory cerebral calcitonin gene-related peptide (CGRP) are mediated by sympathetic noradrenergic efferents acting upon α-adrenergic receptors (170, 222). Sham feeding and electrical vagus nerve stimulation in dogs triggers the release of cholecystokinin (CCK) although this response may be absent in humans (9, 165, 305). Endogenous CCK was shown to enhance PP release in humans during sham feeding (159). Therefore, although peptidergic neurotransmitters are released during vagal stimulation, acetylcholine is believed to be the main neurotransmitter which regulates the cephalic phase.
B. Gastric Phase
Entry of food into the stomach initiates the gastric phase of pancreatic secretion. This phase has been difficult to study in anaesthetized animals because presence of food in the stomach initiates neural reflexes and release of hormones. Therefore, physiological data regarding this phase has been collected by gastric distention induced either by balloon dilation or instillation of inert substances in the antrum. Experiments in which gastric contents were prevented from emptying into the duodenum demonstrated that the gastric phase accounted for approximately 10% of pancreatic secretion. Secretions induced during this phase consist mainly of enzymes with minimal release of bicarbonate suggesting that acinar cells are primarily involved in the induction of this phase (9, 41, 346, 353).
The role of gastrin in this phase of pancreatic secretion remains unclear. Step-wise alkaline distension of the antrum induced graded release of gastrin and pancreatic enzymes (58). However, when exogenous gastrin was administered to dogs the amount required to stimulate exocrine secretion was much greater than normal postprandial gastrin levels, suggesting that gastrin did not have a physiological role (170). These findings are supported by other studies demonstrating that gastrin release is not required for pancreatic enzyme secretion during this phase.
The vagus nerve plays an important role in the gastric phase of pancreatic secretion. Early experiments in anesthetized cats demonstrated that stimulation of the antrum resulted in vagal stimulation of pancreatic amylase release (27). Antral distension in dogs also increased pancreatic secretion by long route vagal pathways (58). An antropancreatic short reflex pathway which is blocked by hexamethonium and atropine also mediates this phase (87). In addition, atropine and vagotomy block the gastric phase providing further evidence that gastric contributions to pancreatic secretion are mediated by vagovagal cholinergic reflexes that originate in the stomach and terminate in the pancreas (183, 353, 354). CCK release plays an important role in antral motility and gastrin release in humans as suggested by sham feeding experiments (159).
In the stomach, pepsin and gastric lipases catabolize proteins and fats into peptides and triglycerides plus fatty acids, respectively, while salivary amylase contributes to the continued digestion of carbohydrates. Peptic digests of proteins are effective in stimulating the intestinal phase (105). Thus when gastric chyme enters the duodenum, it stimulates the intestinal phase of pancreatic secretion. In a clinical setting, surgical procedures that slow the rate of gastric emptying reduce pancreatic secretion (211, 218). Therefore, the rate of gastric emptying regulates the discharge of nutrients into the intestine and consequently the activation of the intestinal phase through neural and hormonal pathways.
C. Intestinal Phase
Digestion of food in the stomach is followed by release of acidic chyme into the duodenum which initiates the intestinal phase of pancreatic secretion. The pancreas has already been primed by cephalic and gastric influences, which enhance blood flow and initiate exocrine secretion. A majority of the pancreatic secretory response (50 - 80%) occurs during the intestinal phase and is regulated by hormonal and neural mechanisms.
The intestinal phase is more easily studied than the gastric phase as food can be instilled directly into the intestinal lumen without concern for gastric emptying. Stimulation of both acinar and ductal cells results in the production of enzyme and bicarbonate secretion. Pancreatic digestive enzyme secretion is stimulated by food molecules such as sodium oleate, monoglycerides, peptides, and amino acids (particularly tryptophan and phenylalanine) (55, 93, 199, 226-228). In the duodenum the high volume of bicarbonate released neutralizes the acidity of gastric chyme, while pancreatic enzymes catabolize partially digested food into molecules that are easily absorbed by intestinal enterocytes.
In the intestinal phase, pancreatic response is regulated primarily by the hormones secretin and CCK, and by neural influences including the enteropancreatic reflex which is mediated by the enteric nervous system and amplifies the pancreatic secretory response. Entry of low pH gastric chyme into the intestine stimulates release of secretin from S cells into the blood (174). The main action of secretin is to stimulate bicarbonate release from pancreatic duct cells, but it also has a direct effect on acinar cells and potentiates enzyme secretion. CCK is released by proteins and fats and their partial digestion products: peptides and fatty acids. Experiments in dogs with chronic pancreatic fistulae have shown that CCK antagonism diminishes pancreatic protein response to a meal and duodenal perfusion suggesting that CCK plays an important role in this phase (179). Similar results were also obtained in humans, where CCK receptor antagonism reduced pancreatic enzyme secretion during the intestinal phase (90, 122).
Cholinergic regulation plays a critical role during this phase of pancreatic secretion. In the absence of secretin, atropine partially inhibits pancreatic bicarbonate secretion stimulated by low pH due to acidic chyme in the duodenum (320, 365). In addition, the amount of bicarbonate produced by infusion of secretin is lower than that released by entry of food into the duodenum suggesting that other factors contribute to meal-stimulated pancreatic bicarbonate secretion (34). Atropine inhibited pancreatic enzyme secretion from 30 -120 minutes following meal ingestion, implicating cholinergic mechanisms (34). Vagovagal enteropancreatic reflexes mediated by M1 and M3 muscarinic receptors and CCK receptors play an important role in the intestinal phase of secretion (316, 318). These vagovagal enteropancreatic reflexes are modulated by input from the dorsal motor nucleus of the vagus projecting into the pancreas. Vagal stimulation activates pancreatic bicarbonate secretion through both cholinergic muscarinic and noncholinergic transmission.
Role of gastric acid
The physiological effects of acid on pancreatic secretion were evaluated by various methods such as diversion of gastric and pancreatic contents with fistulae, and instillation of acidic solutions into the duodenum. Both gastric acid and exogenous HCl are powerful regulators of postprandial pancreatic bicarbonate secretion and their effects are potentiated by intrapancreatic and vagovagal neural pathways as well as by CCK (317).
Intraduodenal infusion of hydrochloric acid elicited a concentration-dependent increase in both the amount of bicarbonate and volume of pancreatic secretion. Secretion was similar to that attained with intravenous infusion of exogenous secretin suggesting that pH changes resulting from entry of acidic contents into the duodenum are important in inducing pancreatic secretion. Administration of a peptone meal of varying pH (pH 1 to 5) produced a maximal secretory response at pH 3.0, which was comparable in magnitude to that obtained with exogenous secretin (63). Acid infusion in both the duodenum and upper jejunum elicited pancreatic secretion suggesting that the proximal small intestine responds to this stimulus (174).
Entry of gastric contents into the duodenum creates an acidic environment with a pH of 2.0 - 3.0 in the initial segment of the duodenum, while the pH of the distal segment remains alkaline (36, 292). This difference in pH is largely due to pancreatic bicarbonate release, which is augmented in large part by gastric acid-induced secretin release from the intestinal mucosa. In conscious rats with gastric and pancreatic fistulae, diversion through a gastric fistula produced a small increase in pancreatic secretion. However, instilling hydrochloric acid into the duodenum with an open gastric fistula augmented pancreatic secretion (24, 99). In addition, pancreatic bicarbonate secretion was much greater when pancreatic juice was diverted from the intestine signifying a correlation between intestinal pH and quantity of pancreatic bicarbonate release (53, 118).
The pancreatic bicarbonate response is dependent on the concentration of free unbuffered hydrogen ions and not on the total load of buffered acid entering the duodenum. Inhibition of gastric acid production by cimetidine (an histamine H2 receptor blocker) or omeprazole (an H+/K+ ATPase inhibitor) substantially reduced the pancreatic bicarbonate response to a meal (24, 243). The pH of a liquid gastric meal also plays a significant role in pancreatic bicarbonate secretion; in cats and dogs, pH > 4.5 resulted in little pancreatic bicarbonate secretion, while at pH <4.0 secretion increased substantially suggesting that a pH threshold of < 4.5 is critical for stimulation of pancreatic secretion (63, 230). This evidence implies that gastric acid is an important regulator of pancreatic bicarbonate secretion which neutralizes the acid to create an alkaline environment optimal for the action of pancreatic enzymes and continued digestion of food.
Role of dietary fat in pancreatic secretion
Dietary fats stimulate pancreatic enzyme and bicarbonate secretion. Intestinal perfusion of monoolein stimulated pancreatic enzyme secretion in humans and this effect was similar in potency to that observed with intravenous CCK injection (213). In contrast, triglycerides administered directly into the duodenum (in the absence of endogenous lipase) were unable to induce pancreatic secretion. However, following lipase digestion of fatty acids, monoglycerides stimulated pancreatic secretion but glycerol was ineffective indicating that fatty acids are the major component of ingested fats that stimulate pancreatic secretion (213, 225).
There is some evidence to suggest that both free and saponified fatty acids induce pancreatic secretion, while other experiments suggest effectiveness only in a micellar form. Secretion has been shown to be dependent on fatty acid chain length, with C4 being least effective and C18 being most effective (65). Although the reason for this difference in potency is not entirely clear, it is not believed to be related to the efficiency of absorption (212). Other studies have demonstrated that intraduodenal administration of propionate (C3) was more effective than oleate (C18) in stimulating acinar cell secretion (253). The reason for the differences between the two studies is not entirely clear but could be species related as these experiments have been performed in humans, rats, and rabbits. Both oleate and neutral fats stimulate bicarbonate and fluid secretion, whereas only neutral fats stimulate pancreatic enzyme secretion. In dogs, oleic acid was shown to potentiate acidified protein-meditated pancreatic enzyme and bicarbonate secretion (80). Fat emulsions given to conscious rats produced a 3-fold increase in pancreatic protein secretion. The route of fat administration also has an impact on pancreatic secretion. Intravenous administration of fat did not produce pancreatic secretion, whereas intraduodenal administration led to elevated protein, bicarbonate, and fluid secretion (258, 329).
Administration of fat emulsions increases plasma CCK and secretin levels. Fat-mediated pancreatic secretion was blocked by proglumide, a CCK receptor antagonist, implicating the importance of CCK in stimulating pancreatic secretion (102). Both C12 and C18 fatty acids augment the effects of secretin-induced bicarbonate secretion (79). In humans, introduction of different concentrations of oleic acid into the duodenum induce pancreatic secretion, although the threshold for CCK stimulation is much lower than for secretin (265). Secretin release is physiologically important since injection of anti-secretin antibodies in conscious rats greatly reduce fat-mediated protein and bicarbonate secretion (106).
A critical fatty acid chain length of C12 was required for CCK release from STC-1 cells, a neuroendocrine tumor cell line. Fatty acids with less than ten carbon atoms did not augment secretion. This dependence on fatty acid chain length is similar to that observed previously for in vivo CCK release in humans. In addition to the fatty acid carbon chain length, a free carboxyl terminus is also important as esterification of the carboxylic terminus abolished CCK secretion, while modification of the methyl terminus had no effect (219-221). Two cell surface receptors have been identified and demonstrated to promote fat-mediated CCK release. Mice with global deletion of GPR40 show partial reduction in CCK secretion following fatty acid administration (204). The recently discovered immunoglobulin-like domain containing receptor (ILDR) is expressed in the duodenum in a subtype of EECs known as I cells, that secrete CCK. ILDR appears to play an essential role in fat-stimulated CCK release as deletion of ILDR in mice completely eliminates fatty acid-stimulated CCK secretion (44).
Thus fats and fatty acids are important regulators of pancreatic secretion. Experimental evidence suggests that the degree and extent of acinar and ductal cell activation may vary depending on the animal species and the route of fat administration.
Contributions of proteins, peptides and amino acids to pancreatic secretion
Studies performed in dogs, rats, and humans have shown that proteins, peptides, and amino acids stimulate pancreatic secretion (325). In rats, proteins such as casein stimulate pancreatic secretion in proportion to their trypsin inhibitor capacity (101). However, in dogs, intact, undigested proteins such as casein, albumin, and gelatin did not stimulate pancreatic secretion, whereas protease digests of these proteins were very effective (226). In contrast, studies in rats suggested that intestinal administration of hydrolyzed casein produced a smaller response than some of the other proteins which potently stimulated pancreatic enzyme secretion, suggesting that the amino acid composition of a protein is relevant in determining the extent of stimulation (101, 200).
Although intravenous infusion of amino acids in humans stimulated pancreatic enzyme and bicarbonate secretion, a mixture of L-amino acids when infused intravenously in dogs was not effective. In contrast to intravenous infusion, intraduodenal delivery of amino acids in dogs induced pancreatic fluid, bicarbonate and protein secretion which was comparable to an elemental diet suggesting the importance of the route of administration on pancreatic secretion (329, 360). Only L-amino acids stimulate pancreatic secretion which is consistent with the overall physiological importance of these stereoisomers. Of all the amino acids, aromatic amino acids such as phenylalanine and tryptophan have the greatest potency (81, 224, 228).
Acidification of amino acid (176, 224) and peptide (81) preparations with hydrochloric acid potentiates the bicarbonate response but pancreatic enzyme secretion is not influenced beyond that observed in the absence of acid. Aromatic amino acids are capable of inducing maximal secretory response as potentiation of pancreatic enzyme secretion is not observed when amino acids or peptides are administered concomitantly with lipid molecules such as oleate or monoolein (80, 213).
The pancreatic secretory response to intraduodenal administration of amino acids appears to be concentration dependent. A minimal concentration of 8 mM is necessary for stimulation by most amino acids (228) although the more potent aromatic amino acids such as tryptophan stimulate secretion at concentrations as low as 3 mM (319). The length of the intestine exposed to amino acids also plays a critical role in pancreatic secretion. In dogs, exposure of the first 10 cm was least effective, while perfusion of the whole intestine produced significant enzyme output (228) suggesting that the pancreatic response was dependent upon the entire load of nutrients, not just their concentration. The majority of stimuli responsible for pancreatic stimulation originate in the proximal small intestine. In humans, amino acids stimulated pancreatic secretion only when perfused into the duodenum and no response was observed upon perfusion in the ileum (68). Therefore, similar to fats, the primary mechanisms that stimulate pancreatic secretion are limited to the proximal regions of the small intestine.
The amount of bicarbonate released by intraluminal administration of tryptophan is similar to that produced by maximal doses of exogenously infused CCK indicating that release of CCK by tryptophan leads to pancreatic secretion (57, 213, 226). Similarly, intraduodenal administration of liver extracts in dogs mediated CCK release along with pancreatic enzyme and bicarbonate secretion, both of which were blocked by CCK receptor antagonists (245). Bile acids released from the gallbladder can significantly inhibit pancreatic stimulation induced by intraluminal amino acids. This inhibition of pancreatic secretion by bile acids appears to be due to inhibition of CCK release and serves as a feedback mechanism in regulating pancreatic and gallbladder function (213). By using a sensitive bioassay for CCK measurement, it was shown that one of the pathways by which proteins stimulate CCK release is by their ability to inhibit intraluminal trypsin activity (200). Another mechanism by which aromatic amino acids mediate CCK release is by activation of the calcium sensing receptor (CaSR), a known nutrient sensor (123, 205, 252, 351). In addition to stimulating the release of hormones such as CCK and secretin, amino acids also activate cholinergic neural mechanisms which regulate pancreatic bicarbonate secretion (319).
Role of bile and bile acids in pancreatic secretion
Bile is produced by hepatocytes as a complex mixture of bile acids, cholesterol, and organic molecules. It is stored and concentrated in the gall bladder and released into the duodenum upon entry of chyme. Bile acids such as cholate, deoxycholate, and chenodeoxycholate are conjugated with glycine or taurine amino acids which increase their solubility. In the intestine, bile acids assist in the emulsification and absorption of fatty acids, monoacylglycerols, and lipids and stimulate lipolysis by facilitating binding of pancreatic lipase with its co-lipase.
Under basal conditions, intraduodenal administration of physiological concentrations of bile or the bile salt sodium taurocholate, elevated plasma secretin and stimulated pancreatic fluid secretion in cats (109, 110). Secretin was released only in response to perfusion of sodium taurocholate in the duodenum. Perfusion in the upper jejunum produced a significantly diminished pancreatic response, while no response was observed upon ileal perfusion (112). Pancreatic fluid secretion was stimulated by the free ionized form of taurocholate and was not dependent on its detergent properties (103). In humans, infusion of bovine bile augmented secretin release along with pancreatic exocrine secretions of fluid, bicarbonate, and enzymes (266, 267).
In addition to secretin, infusion of bovine bile and bile acids in humans and dogs was shown to stimulate the release several hormones and neuropeptides such as CCK, neurotensin, VIP, gastric inhibitory peptide (GIP), PP, and somatostatin (38, 47, 287, 289). Fluid and bicarbonate release was enhanced when elevated levels of VIP were present in the plasma, suggesting that bile activates peptidergic nerves resulting in pancreatic secretion. Additionally, cholinergic mechanisms are also important as atropine blocked bile- and taurocholate-stimulated exocrine pancreatic secretion (289). The composition of bile is important in mechanisms regulating this secretory response as some differences in hydrokinetic and ecbolic responses were observed with administration of bile versus various bile acids (286).
However, a stimulatory effect of bile acids on pancreatic fluid secretion was not observed in the presence of digestive intraluminal contents (31). In some studies, where bile acids were administered concomitantly with amino acids or fat, an inhibition of pancreatic enzyme secretion was observed. The mechanism underlying this observation is not completely understood, although it is possible that bile acids inhibit CCK release by a negative feedback mechanism which helps to relax and refill the gallbladder (26, 181, 213, 261). Chemical sequestration of bile acids in dogs augmented the release of CCK and pancreatic enzyme secretion in response to amino acids and addition of taurocholate reversed this effect (94). Long term diversion of bile in dogs also augmented basal and oleate-stimulated pancreatic fluid, bicarbonate, and enzyme secretion along with plasma CCK levels, further supporting the role of bile acids in inhibiting CCK release (333).
Other studies have shown that the bile salt chenodeoxycholate when infused in humans, inhibited bombesin- and CCK-stimulated gallbladder emptying along with elevation of plasma CCK levels. These results led the authors to hypothesize that chenodeoxycholate, by a yet unknown mechanism, reduced the sensitivity of the gallbladder to stimulation by bombesin and CCK (340).
In contrast to many species including mice and humans, rats do not possess a gallbladder and multiple pancreatic ducts join the lower end of the common bile duct. In rats, diversion of bile and pancreatic juice stimulates the release of pancreatic enzymes. This augmentation of enzyme secretion has been suggested to compensate for the increased degradation of proteolytic enzymes in the absence of bile. Thus exocrine secretion in rats is regulated by a luminal feedback mechanism (98, 236). Additional experiments have shown that certain bile salts stimulate bicarbonate secretion via CCK release whereas other bile salts inhibit exocrine secretion (234, 237, 238). Two inhibitory mechanisms have been proposed – one dependent on the stabilization of luminal proteases and the other independent of protease activity (239). Stimulation of pancreatic fluid secretion in anesthetized rats has been demonstrated to be mediated by taurocholate induced transcription of Na+/K+/2Cl- cotransporter, which plays a key role in regulating the entry of Cl- from the basolateral surface of acinar cells.
The physiological role of bile and bile salts in regulating pancreatic secretions is not completely understood and appears to be dependent on multiple factors, including the chemical properties of bile salts, the animal model being evaluated, and prandial status of the animal being studied (288).
D. Absorbed Nutrient Phase
Once nutrients are absorbed from the intestinal lumen, they can directly stimulate pancreatic secretion as part of l the absorbed nutrient phase. Nutrients can either directly stimulate pancreatic acinar cells, or they may indirectly activate hormonal and neural pathways to further regulate exocrine secretion. In contrast to intraduodenal administration of amino acids which produces large increases in pancreatic secretion (178, 229, 291) intravenous administration of amino acids stimulates trypsin and chymotrypsin secretion, but not lipase or amylase (91). Less is known about lipids and glucose although they appear to have little if any direct effect on pancreatic secretion (198, 258).
E. Feedback regulation of pancreatic secretion
The concept of feedback regulation of pancreatic secretion emanated from a series of studies demonstrating that (1) instillation of trypsin inhibitor into the upper small intestine or (2) surgical diversion of the bile-pancreatic duct removing bile and pancreatic juice from the duodenum of rats stimulated pancreatic enzyme secretion (100). Conversely, infusion of trypsin into the duodenum during bile-pancreatic juice diversion suppressed pancreatic enzyme release. Thus, the protease concentration in the upper small intestine appears to be intimately linked to pancreatic secretion through a negative feedback system in which active proteases within the duodenum limit pancreatic secretion but reduced protease activity stimulates pancreatic secretion. When assays for CCK became available, it was shown that CCK mediated the effects of proteases on pancreatic secretion (197) through protease-sensitive CCK releasing factors (121, 328) (see Figure 1). In the absence of proteases, CCK releasing factor can stimulate CCK cells, but in the presence of proteases, the releasing factors are inactivated and CCK secretion is low. Negative feedback regulation of pancreatic secretion has been shown to exist in many species although other proteases such as elastase may be more important in regulating pancreatic secretion in humans.
Figure 1. Feedback regulation of pancreatic exocrine secretion is mediated by positive and negative mechanisms. Positive feedback: Monitor peptide is secreted by acinar cells and directly stimulates CCK cells in the small intestine and amplifies pancreatic secretion once it has been initiated. Negative feedback: Trypsin-sensitive CCK releasing factors are produced by the intestine and stimulate CCK secretion when trypsin is temporarily consumed by ingested protein or other trypsin “inhibitors”.
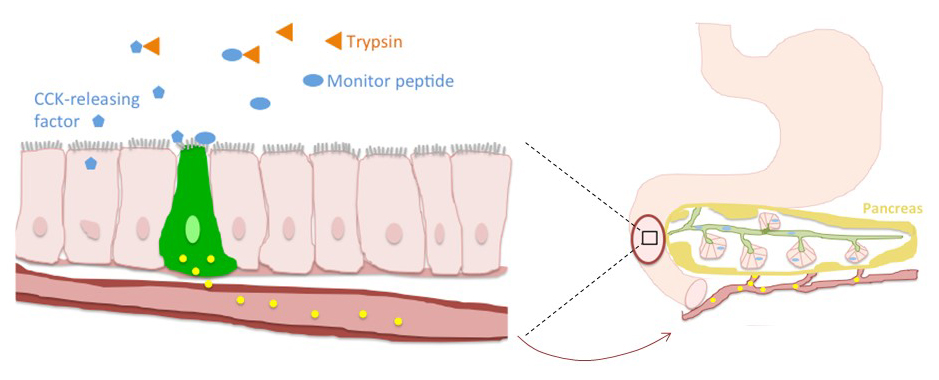
Pancreatic exocrine secretion is also influenced through a positive feedback mechanism. Monitor peptide is a 61 amino acid peptide produced by pancreatic acinar cells and possessing CCK releasing activity. Although monitor peptide has modest trypsin inhibitor capability, its ability to stimulate CCK is independent of this action because monitor peptide can directly stimulate CCK secretion from isolated CCK cells in vitro (32, 201). Monitor peptide is secreted in pancreatic juice, therefore, it does not stimulate CCK secretion unless pancreatic secretion is underway. Thus, monitor peptide cannot account for the increase in CCK in during bile-pancreatic juice diversion, but it may serve to reinforce pancreatic secretion once the process has been initiated.
III. PANCREATIC EXOCRINE SECRETION
The exocrine pancreas delivers its secretions of digestive enzymes, fluid, and bicarbonate ions to the duodenum following ingestion of food. The pancreas is composed of both endocrine and exocrine components. The endocrine pancreas is comprised of α, β, δ, ε, and PP (F) cells, which are located in the islets of Langerhans. These specialized cells secrete the hormones glucagon, insulin, somatostatin, ghrelin, amylin, and pancreatic polypeptide into the blood, which exert endocrine and paracrine actions within the pancreas. Ninety percent of the pancreas is composed of acinar cells which secrete digestive enzymes such as trypsin, chymotrypsin, and amylase for digestion of food in the small intestine. The acinar cells are triangular in shape and arranged in clusters with the apex of the cell opening into a centrally located terminal duct. The terminal or intercalated ducts merge to form interlobular ducts, which in turn congregate to form the main pancreatic duct. The pancreatic duct delivers exocrine secretions into the duodenum. The ductal cells secrete fluid and bicarbonate ions, which neutralize acinar cell secretions, as well as the acidic gastric contents entering the duodenum (115). The pancreas is heavily innervated by sympathetic and parasympathetic peripheral nerves and contains a dense network of blood vessels which regulate blood flow and modulate pancreatic secretion.
Pancreatic exocrine secretion is a highly integrated process mediated by neural and hormonal signals arising from the gut as well as by factors secreted by other tissues and hormones released from pancreatic islets. The secretory pathways can be stimulatory or inhibitory in nature, and represent a highly regulated system that responds to ingestive signals. The agents that modulate pancreatic exocrine secretion are discussed below (Table 1).
A. Neural Mechanisms
The pancreas is innervated by parasympathetic nerve fibers, postganglionic sympathetic neurons, as well as a network of intrapancreatic nerves. Together these nerves regulate pancreatic exocrine function by releasing neurotransmitters such as acetylcholine, serotonin and neuropeptides such as VIP, GRP, and neuropeptide Y (NPY). The pancreatic ganglia receive input from pre- and post-ganglionic nerve fibers and regulate exocrine and endocrine secretion.
Intrapancreatic postganglionic neurons are activated by central input during the cephalic phase and by vagovagal responses initiated during the gastric and intestinal phases of stimulation. They stimulate enzyme and bicarbonate secretion primarily by releasing acetylcholine, which activates muscarinic receptors located on acinar and duct cells.
Vagal Innervation
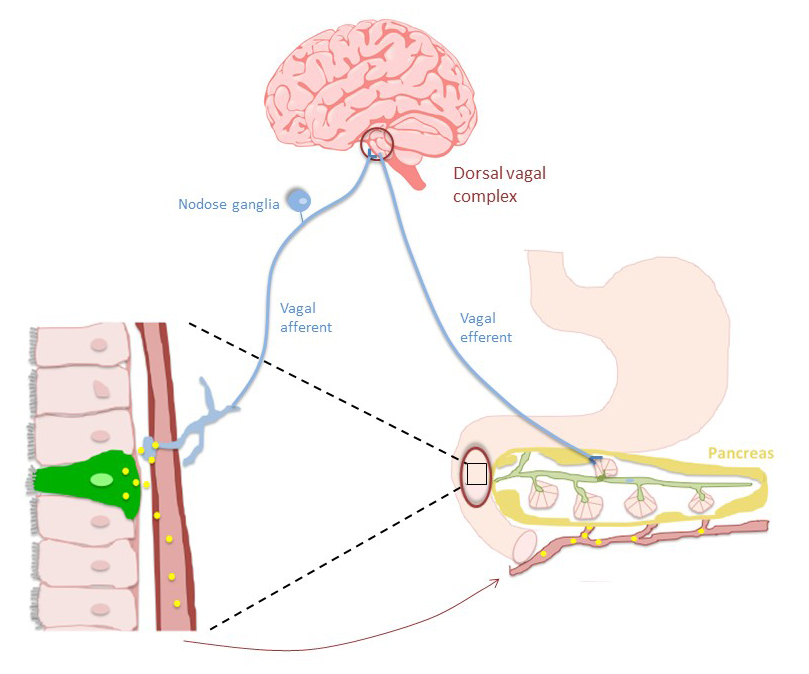
The dorsal vagal complex in the brainstem is comprised of the nucleus of the solitary tract (NTS) and the dorsal motor nucleus of the vagus (DMV) and exerts parasympathetic control on pancreatic secretion. Information relayed by sensory vagal afferent nerves innervating the pancreas is first processed in the NTS, which then projects onto the preganglionic motor neurons of the DMV. The DMV receives inputs from other regions of the brain such as the hypothalamus and from numerous hormones and neuropeptides through the afferent limb of the vagus nerve (12).
Parasympathetic preganglionic efferent vagal nerves innervating the pancreas originate primarily from the DMV and terminate in the pancreatic ganglion. Electrical and chemical stimulation of the DMV induces rapid pancreatic secretion, and this response is inhibited by vagotomy or blockade of muscarinic receptors by atropine (251). It has been suggested that vagal cholinergic neurons mediate pancreatic secretion during low loads of intestinal stimulants whereas hormones mediate the response during high loads of intestinal stimuli (257, 318).
CCK affects pancreatic secretion through both a direct effect on pancreatic acinar cells and an indirect effect on the vagus nerve (Figure 2). However, the effects on the vagus nerve are complex and the firing response of neurons in the DMV appears to be dictated by their spatial location. In one study, neurons in the caudal region were activated, those in the rostral region were unaffected, while neurons in the intermediate region were inhibited by a direct action of CCK (250). Although it is not fully understood, it appears that CCK’s effects on the vagus nerve influences the overall pancreatic secretory response.
The exocrine pancreas is regulated directly by the vagus. Studies with muscarinic receptor knockout mice demonstrated that both M1 and M3 receptors mediate amylase release from dispersed acini. It is likely that M3 receptors are more relevant physiologically since the level of M3 receptor expression was significantly higher in acinar cells (92) and M1 receptors were found to have only a minor effect on bicarbonate secretion in conscious dogs (339). The vagus nerve also possesses group II metabotropic glutamate receptors that couple primarily to Gi/o. These receptors are located on excitatory and inhibitory pre-synaptic terminals of pancreas-projecting DMV neurons (11) that are also activated by CCK and pancreatic polypeptide. Thus, in addition to γ-amino butyric acid, glutamate also modulates pancreatic exocrine secretion through distinct vagal neurons.
Figure 2. CCK stimulates pancreatic secretion through hormonal and neuronal pathways. CCK is released from I cells of the small intestine and diffuses into the blood stream where it is carried to the pancreas. CCK binds to receptors on acinar cells to stimulate pancreatic enzyme secretion. Secreted CCK also diffuses through the paracellular space and binds to CCK1 bearing nerves in the submucosa. Vagal afferent signals are integrated in the dorsal vagal complex which also receives signals from other regions of the brain (e.g., hypothalamus). Vagal efferent fibers transmit cholinergic signals to the pancreas to stimulate pancreatic secretion.
Vasoactive Intestinal Peptide (VIP)
Vasoactive Intestinal Peptide is a 28 amino acid neuropeptide that is found throughout the body. Immunocytochemical evidence suggests that VIP is localized in pancreatic nerve fibers and functions as a vagal neurotransmitter. In the chick, VIP immunoreactive nerve endings are found in close proximity to acinar cells and epithelial cells of arterioles. Small clear vesicles were present in VIP-positive nerves indicating that these neurons are were cholinergic in nature (124). In normal human pancreas, autonomic ganglia receive an abundant supply of VIP-positive fiber plexi, and VIP-positive nerves and appeared to innervate acinar cells, ducts, and blood vessels (215). After atropine treatment, electrical stimulation of the vagus still increased bicarbonate secretion concurrent with detection of VIP in pancreatic venous effluent suggesting that VIP release is coupled with bicarbonate secretion (75). The effects of VIP are especially prominent in the pig as perfusion of the pancreas with VIP antibodies inhibited fluid and bicarbonate secretion, and treatment of rats with a VIP antagonist reduced bicarbonate secretion concomitant with vasodepression further supporting a direct relationship (126, 352). High and low affinity VIP receptors have been identified on pancreatic acinar membranes. The high affinity receptors are coupled to cAMP-mediated amylase release, while activation of low affinity receptors did not cause cAMP elevation or amylase release, suggesting that only high affinity receptors are important in protein secretion (25). These effects of VIP were attenuated by somatostatin and galanin, which reduced VIP-mediated fluid and protein output (129). One of the main functions of VIP appears to be increasing blood flow by vasodilation, and as a result its effects on pancreatic secretion independent of blood flow in the pancreas are difficult to evaluate (10, 180).
Gastrin Releasing Peptide (GRP)
Gastrin releasing peptide is a 27 amino acid neuropeptide that is present in post-ganglionic vagal afferents and has been detected in neurons innervating the feline, porcine, rodent, and human pancreas (242). Receptors responsive to GRP have been identified in rat pancreatic membranes and cancer cells where they mediate enzyme secretion (108, 156). In the cat, GRP is present in intrapancreatic ganglia, acinar and stromal regions, and occasionally on the vasculature and ducts (56). In humans, the pattern of GRP expression was similar to that of VIP; GRP was localized on nerve fibers in proximity to pancreatic acini, capillaries, ductules, and arterial walls (312). Several studies in different species have demonstrated that GRP modulates exocrine secretion. Vagal stimulation of porcine pancreas resulted in GRP release which enhanced pancreatic exocrine secretion (167). In isolated perfused rat pancreatic preparations, electrical field-stimulated GRP release potentiated secretin-mediated fluid and amylase secretion through a non-cholinergic pathway (277). The effects of GRP on rat pancreatic exocrine secretion were enhanced by γ-amino butyric acid (279). Neuromedin C, a decapeptide of GRP, also enhanced pancreatic secretion by direct action on canine acinar cells as well as indirectly by stimulating CCK release (134, 135). However, since bombesin (GRP analog) does not stimulate pancreatic secretion in dogs, it appears that its effects may be dependent on the species being evaluated (177).
Other Peptide Neurotransmitters
Immunohistochemical staining has revealed that the neuropeptides listed below are present in pancreatic nerves and their functional significance and ability to regulate pancreatic secretion has been demonstrated by in vitro and/or in vivo studies.
- PACAP: Pituitary adenylate cyclase-activating polypeptide (PACAP) has been identified in nerve fibers and intrapancreatic ganglion in rodents (84, 96). PACAP has been shown to evoke bicarbonate and enzyme secretion from the pancreas albeit with a slower time course than VIP (8, 352). In the acinar cell line AR42J, PACAP activated phospholipase C, which led to elevation of intracellular Ca2+ and amylase release (15).
- Neurotensin: Neurotensin is a 13 amino acid neuropeptide that is widely expressed in the central nervous system and is also present in pancreatic nerves. It stimulates amylase secretion and its effects are potentiated by carbachol (a cholinergic agonist), secretin, and caerulein (a CCK analog) (13, 78). Other studies demonstrated that neurotensin stimulates bicarbonate, but not protein secretion in dogs and may act indirectly by stimulating dopamine release (142).
- Substance P: Substance P is expressed in periductal nerves in the guinea pig pancreas and inhibits ductal bicarbonate secretion by modulating neurokinin 2 and 3 receptors (116, 162, 169). It enhanced caerulein-stimulated enzyme secretion in isolated perfused pancreas as well as in anesthetized rodents (158).
- CGRP: CGRP is a 37-amino acid peptide that is present in central and peripheral neurons. The effect of CGRP on exocrine secretion is not clear and may be species specific. Interaction of CGRP with receptors on guinea pig acinar cells led to amylase release, although its effect was not as potent as VIP (309). In rat acinar cell preparations, CGRP inhibited amylase release by a mechanism involving cholinergic (muscarinic) neural pathways (37).
- NPY: NPY, a 36 residue peptide, is expressed in intrapancreatic ganglia and nerve fibers that surround exocrine pancreatic tissue (310). NPY inhibited CCK- and vagally-mediated amylase secretion from intact pancreas and lobules, but not from dispersed acini, suggesting that its actions were mediated by neurons innervating the exocrine pancreas (246). Other evidence suggests that NPY plays at best only a modest role in pancreatic exocrine secretion (128).
- CCK: The presence of CCK in intrapancreatic nerves has led to the suggestion that it may serve a dual role as neurotransmitter and hormone in pancreatic secretion. However, CCK could not be detected in the venous effluent of isolated perfused porcine pancreas after vagal stimulation following a meal (126) suggesting that synaptic release of CCK does not occur within the pancreas. Therefore, the role of CCK as a neurotransmitter in pancreatic secretion merits further investigation.
- Peptide Histidine Isoleucine: Peptide histidine isoleucine (PHI) is a 27 amino acid peptide with an N-terminal histidine and C-terminal isoleucine, and is derived from the same precursor molecule as VIP. It has been reported in pancreatic nerves and ganglia and stimulates fluid and bicarbonate secretion in a cAMP-dependent fashion (143, 310).
Adrenergic Nerves
Compared to cholinergic stimulation, adrenergic nerves play a relatively minor role in pancreatic exocrine secretion. Catecholamine-containing nerves are found in the celiac ganglion, and extend to intrapancreatic ganglia, blood vessels, ducts, and islets (186). Epinephrine and norepinephrine (NE) evoked amylase release from superfused rat pancreatic preparations, similar to that induced by electrical stimulation in the presence of cholinergic blockade (323). This process is dependent on elevated intracellular Ca2+ and inhibited by propranolol, suggesting that β-adrenergic receptors are involved (281). Catecholamines also interact with α-adrenergic receptors expressed in pancreatic acini and inhibit amylase secretion (348). NPY is coexpressed with NE in some nerve fibers, and stimulation of splanchnic nerves leads to the release of NPY and NE (42, 310). Infusion of PACAP into the pancreatoduodenal artery enhanced release of NE after electrical stimulation of nerves. However, the physiological relevance of this observation is not clear (363).
Celiac denervation reduces pancreatic secretion by ~70% while increasing blood flow by 350%. This dissonance presumably occurs by the disruption of stimulatory fibers and sympathetic fibers that maintain tonic constriction of pancreatic vessels (166). The effect of adrenergic transmitters on pancreatic secretion has been difficult to discern due to the wide-ranging effects of norepinephrine on multiple processes including blood pressure, blood flow, neural reflexes, and release of hormones. Even though high concentrations of norepinephrine have been found in rabbit pancreatic ganglia, ducts, and blood vessels, its effects are controversial (364). Norepinephrine has been reported to stimulate, inhibit, or have no effect on pancreatic secretion (14, 46, 64, 89, 186, 193, 326). Unfortunately α- and β- adrenergic receptor agonists and antagonists have not provided information that could be used to delineate mechanisms important in adrenergic regulation of pancreatic secretion (45, 66).
Dopamine
Dopamine was detected in pancreatic ducts and ampullae and dopamine β-hydroxylase (DBH) -positive fibers were identified along the vasculature, ducts, and ganglia suggesting it may play some role in pancreatic secretion (209, 364). There is conflicting evidence regarding the role of dopamine in pancreatic secretion. Dopamine stimulates pancreatic secretion in anesthetized dogs and rats although the effect is negligible in conscious animals (19, 60, 64, 89, 138, 141). Other data suggest that the secretory response to dopamine differs between dogs, cats, rabbits and rats, and species specific effects must be taken into consideration when evaluating its effects (114).
Serotonin
Like dopamine, serotonin is present in pancreatic ducts and ampullae. Autoradiography of tissue sections after tritiated serotonin uptake demonstrated the presence of serotonergic innervation of blood vessels, but not exocrine parenchyma in rats, suggesting a limited role in pancreatic secretion (168, 364). However, phenylbiguanide, a 5-HT3 receptor agonist, activated preganglionic neurons located in the caudal DMV, inhibited those in the intermediate DMV, and had no effect on rostral DMV neurons, suggesting complex spatial regulation of pancreatic vagal neurons by serotonin (250). Intraduodenal infusion of melatonin (a serotonin derivative) increased pancreatic amylase secretion, while pretreatment with 5-HT2 serotonin antagonist ketanserin or the 5-HT3 antagonist MDL72222 decreased amylase release. Serotonin-induced amylase release was blocked by bilateral vagotomy supporting a role for serotonergic mechanisms on exocrine secretion (255).
Nitric Oxide (NO)
Nitric oxide is a gaseous signaling molecule that is synthesized by NO synthase (NOS) from L-arginine in the presence of nicotinamide adenine dinucleotide hydrogen phosphate (NADPH). It is a potent vasodilator and modulates secretory activity as well as pancreatic blood flow in the pancreas (49). Because it is not practical to directly measure NO in biological tissues, the presence of NO has been identified by expression of NOS or histochemistry of NADPH diaphorase (NADPH-d), since NOS and NADPH-d colocalize in neurons of the peripheral and central nervous systems. The actions of NO in tissues have been identified by the use of NO donors, NOS inhibitors, and agents that inactivate (e.g. superoxide-generating compounds) or stabilize NO (e.g., superoxide dismutase). Unlike ligands that signal through cell surface receptors, NO penetrates cells and activates guanylate cyclase to generate the second messenger cGMP (361).
Immunostaining of pancreas from a wide range of mammals (mouse, rat, hamster, guinea-pig, cat and man) indicates that NOS is expressed in the cell bodies of intrapancreatic ganglia, interlobular nerve fibers, and along blood vessels. VIP is sometimes co-expressed with NOS in ganglia and nerve fibers. These studies suggest that NO is important in pancreatic exocrine secretion (71). In newborn guinea pig, nitrergic neurons were present primarily in the head and body of the pancreas, along blood vessels, the main pancreatic duct, and in association with pancreatic acini (206). These nerves also immunostained with antibodies against NPY, VIP, and DBH indicating complex co-regulation of pancreatic secretion by various neurotransmitters.
In rat pancreas, the NO donor sodium nitroprusside and cGMP analog 8-bromo cGMP, inhibited basal and vagal amylase secretion through a Ca2+-dependent mechanism (73, 362). The G protein-coupled receptor, protease-activated receptor-2 (PAR-2) modulates NO-mediated amylase release in mice, and inhibition of NOS abolished PAR-2 mediated amylase release suggesting that the effects of NO may be mediated by neuronal release of a PAR-2 agonist. Ablation of sensory nerves by capsaicin did not affect PAR-2 mediated amylase release, suggesting that TRPV1-expressing sensory vagal fibers are not involved in this pathway (160). Analysis of the effects of NO on pancreatic secretions in pigs support the findings that NO is essential for pancreatic fluid and amylase secretion mediated by the vagus nerve (130). Thus, in addition to maintaining the vascular tone, nitrergic nerves play an important role in pancreatic fluid and amylase release.
B. Stimulatory Hormones
Cholecystokinin (CCK)
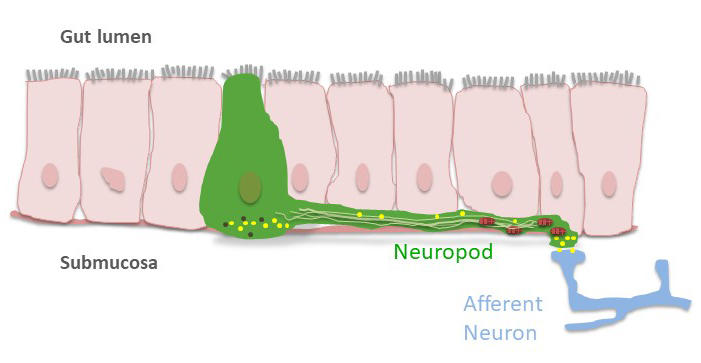
CCK is released from specialized enteroendocrine cells (I cells) located mainly in the upper small intestine. Using confocal microscopy to examine the intestine of transgenic CCK-enhanced green fluorescent protein mice (Cck-eGFP), it was noted that enteroendocrine cells (EECs) possessed basal processes that extended to other cells (43) (Figure 3). In addition to hormones like CCK, it was found that these cells possessed many neuron-like properties including neurotransmitters and synaptic proteins. Their basal extensions, called neuropods, were filled with neurofilaments (29) and connected to nerves (30). Thus, synaptic connections between EECs and nerves, including the vagus nerve (155) provide a way for EECs to sense gut contents and directly regulate neural function.
Figure 3. Model of an enteroendocrine cell (EEC) with a neuropod. EECs (green) are sensory cells that reside in the intestinal mucosa. Some EECs possess neuropods that contain neurofilaments (light green), synaptic proteins, mitochondria (red), and neurotransmitters (yellow) and synapse with neurons in the submucosa. In this manner, EECs may sense intraluminal contents such as nutrients or bacteria and signal directly to nerves, including the vagus nerve.
The major stimulants of CCK release are dietary fats and proteins. In the rat, intraluminal proteases via an active feedback system participate in the release of a putative intestinal CCK releasing factor (e.g., LCRF) which in turn causes CCK secretion (121, 328). Various molecular forms of CCK, ranging in size from CCK-8 [CCK-(26-33)-NH2] to CCK-58, have been described in dogs, rats, and humans (70, 74, 344). CCK-58, was determined to be the predominant peptide in dogs and humans, and the only form detected in rats after employing CCK isolation techniques that prevented degradation of CCK in blood (285). The actions of CCK-8 and CCK-58 appear to be functionally identical suggesting that CCK-8 retains the biological activity ascribed to this hormone (54). CCK is post-translationally modified and has an amidated C-terminus. A sulfated tyrosine residue in CCK-8 is important for its biological actions including exocrine secretion (149, 298). C-terminal amidation is critical for binding of CCK to its receptors and removal of the amide group decreases CCK activity. Other studies have reported that deamidation and desulfation, do not significantly impair the ability of CCK to stimulate amylase release and these discrepancies may arise from differences between species. Shorter forms of CCK such as the tetrapeptide CCK-4, are generally much less effective in mediating exocrine secretion than CCK-8, while longer forms of CCK, such as CCK-33 are equally effective (69, 91, 264, 268).
CCK mediates its hormonal effects through two G-coupled protein receptors, CCK-1 and CCK-2, previously known as CCK-A and CCK-B, respectively. The contribution of these receptors to pancreatic secretion has been evaluated in order to delineate the molecular mechanisms of CCK action. CCK receptors have been proposed to exist in two states, a high affinity (picomolar) but low capacity state, and a low affinity (nanomolar) but high capacity state (336, 347). Autoradiography of pancreatic membranes incubated with radioiodinated CCK-8 demonstrated that CCK-1 receptors are highly expressed in rat pancreas, while CCK-2 receptors are less abundant. CCK-1 receptors appear to modulate pancreatic secretion as oral administration of loxiglumide, a potent CCK-1 receptor antagonist, reduces protein and fluid output in rats (139). Similarly, caerulein-induced pancreatic amylase release was blocked by CCK-1 receptor antagonists (104). CCK-8 did not induce amylase release in CCK-1 receptor knockout mice confirming that CCK-1 receptors are critical for CCK-mediated protein secretion (182). Since bicarbonate secretion was not observed from dispersed acinar cells this effect is not believed to be regulated by CCK receptors (334). Pancreatic responses in CCK-2 receptor knockout mice were similar to wild type mice suggesting that CCK-2 is not important for amylase release, although it may be involved in augmenting vascular flow (104, 241). In pigs, where a majority of receptors are of the CCK-2 subtype, acinar cells demonstrated a low responsivity to CCK and did not secrete amylase in response to caerulein or a CCK-1 agonist (244). In humans, the actions of CCK on pancreatic secretion have been variously reported. Infusion of CCK, caerulein, and secretin in the presence of amino acids substantially increased output of fluid, bicarbonate, and enzyme (172, 332). Similar to porcine pancreas, CCK-2 is the major CCK receptor subtype expressed in human pancreas, although interestingly CCK-1 receptor antagonists are able to inhibit amylase secretion (3, 283, 335). In dispersed human acini which responded to carbamylcholine and neuromedin C, CCK did not stimulate amylase release presumably because of a paucity of cellular membrane receptors. It has been proposed that the effects of CCK on human pancreatic secretion are mediated through CCK-1 receptors on nerves which innervate the pancreas (150, 240). However, recent data demonstrated that application of physiologic concentrations of CCK-8 and CCK-58 to human acinar cells produced intracellular Ca2+ oscillations and normal exocytosis of pancreatic enzyme, suggesting that functional CCK receptors are expressed on human pancreas (249). By visualizing Ca2+ oscillations in a unique slice preparation of human pancreas obtained from cancer resections, it has been unequivocally demonstrated that CCK-8 stimulates pancreatic secretions through direct action on CCK-1 receptors expressed on acinar cells and does not require a neuronal component (196). Thus, it appears that CCK receptors are expressed on acinar cells of both human and rodent pancreas, and the differences between the two may not be as great as previously predicted.
Recently it was shown that rat and human pancreatic stellate cells express CCK receptors and secrete acetylcholine in response to CCK stimulation. This source of acetylcholine was sufficient to stimulate amylase release from acinar cells. Pancreatic stellate cells may represent a previously unrecognized intrapancreatic pathway regulating CCK-induced pancreatic exocrine secretion (282).
Several hormones and neuropeptides regulate CCK-mediated exocrine secretion. Locally, insulin has been shown to influence exocrine secretion. Intra-arterial infusion of canine pancreas with anti-insulin antibodies, prevented CCK release as well as secretin-mediated protein and fluid secretion from canine pancreas (188). Secretin potentiated, as well as attenuated, CCK-mediated amylase secretion by the inositol signaling pathway while VIP enhanced CCK-mediated enzyme secretion (39, 40). Peptide YY (PYY), PP, and somatostatin also inhibited CCK-mediated protein secretion and their effects are discussed later in this review.
The mechanism of CCK-induced amylase secretion, involves transient elevation in intracellular Ca2+ (270). It also requires phospholipase C activation and generation of second messengers inositol trisphosphate and diacylglycerol (272). In some instances, CCK activates secretion by elevation of cAMP, as 8-bromo-cAMP and a phosphodiesterase inhibitor augmented CCK-mediated amylase release (39). Heterotrimeric G proteins Gα13 and Gαq through downstream interactions with small GTP binding proteins RhoA and Rac1 regulate actin cytoskeleton reorganization which is required for exocytosis (293, 359).
Secretin
Secretin is a 27 amino acid hormone released by S cells of the small intestine (358). Secretin release is stimulated during the intestinal phase upon entry of gastric acid and ingested fatty acids into the duodenum (76). It augments fluid and bicarbonate secretion and is one of the most potent stimulators of pancreatic secretion (48). Examination of pancreatic ultrastructure shortly after secretin injection revealed that fluid is secreted by duct as well as acinar cells (28).
Large increases in pancreatic fluid and bicarbonate secretion have been demonstrated with secretin infusions as low as 1-2.8 pmol/kg∙hr (21, 107, 304, 366). In humans bolus injections of secretin as low as 0.125 pmol/kg stimulated fluid and bicarbonate secretion (154, 303). Although secretin is believed to be the single most powerful stimulator of pancreatic bicarbonate secretion, infusion of exogenous secretin equivalent to postprandial blood levels only produced 10% of the maximal pancreatic bicarbonate secretory response suggesting that other hormones and neurotransmitters play important roles in postprandial pancreatic bicarbonate secretion in humans (303, 304). Secretin receptors have been localized in acinar and duct cells in the rat pancreas (345).
Secretin stimulates the release of fluid and bicarbonate, and to a lesser extent, protein from acinar cells by a cholinergic mechanism. Perfusion of acetic and lactic acids in the duodenum of anesthetized rats increased fluid and protein output from the pancreas concomitant with elevation of plasma secretin levels. In addition, treatment of rats with atropine decreased plasma secretin levels and inhibited fluid (but not protein) secretion, indicating that only fluid secretion is dependent on cholinergic input (299). Electrical field stimulation of isolated perfused rat pancreas demonstrated that secretin-mediated exocrine secretion was sensitive to tetrodotoxin and atropine blockade, further suggesting cholinergic regulation. Nicotinic acetylcholine receptors are not involved in this mechanism as hexamethonium did not exert an inhibitory effect on pancreatic secretion (278).
Both cAMP-dependent and independent pathways contribute to secretin-mediated exocrine secretion. Interaction of secretin with its receptor induced a 3-4 fold increase in adenylate cyclase activity which was abolished in the presence of secretin antagonists (223). Secretin did not stimulate pancreatic fluid release or elevate acinar cell cAMP levels in secretin receptor knockout mice (300). Exocrine protein secretion by secretin was associated with phospholipase C activation in one report (341). At secretin concentrations >10-8 M, inositol trisphosphate and diacylglycerol were generated in acinar cells, which caused release of Ca2+ from intracellular stores and activated protein kinase C. However, not all investigators have observed this effect.
Several hormones and peptides modulate the effects of secretin on pancreatic secretion. CCK augmented secretin-induced pancreatic fluid and protein output by stimulating acetylcholine release and this effect was blocked by atropine or by dispersion of acini (7, 327). Venous drainage from pancreatic islets bathes the exocrine pancreas with high concentrations of islet hormones. Several of these hormones have potent effects on pancreatic secretion. Insulin enhances secretin-mediated fluid and protein secretion through an ouabain-sensitive Na+,K+-ATPase while glucagon inhibits secretin-stimulated release of fluid and protein (113). Addition of anti-somatostatin antibodies increased secretion from perfused rat pancreas implying that somatostatin inhibits secretin-induced fluid and enzyme secretion (113, 278).
Glucagon-like peptide 1 (GLP-1)
GLP-1 is a 29 amino acid peptide that is produced by enteroendocrine cells in the distal small intestine ((247). Until recently the role of GLP-1 in pancreatic exocrine secretion was not clear (356). Using real-time PCR it has now been shown that GLP-1 receptor (GLP-1R) is expressed in both pancreatic acinar cells and islets. GLP-1 stimulated amylase release from wild-type but not GLP-1R knockout mice through a cAMP dependent mechanism (136). These results have been supported by Satoh et al., (302) who showed that when isolated rat pancreatic acini were stimulated by GLP, amylase was released in conjunction with phosphorylation of myristoylated alanine-rich C kinase substrate (MARCKS) protein. Inhibition of MARCKS phosphorylation prevented GLP-1 mediated amylase secretion, suggesting it was a critical step in this signaling pathway.
Atrial Natriuretic Factor (ANF) and C-Natriuretic Peptide (CNP)
Atrial natriuretic factor is a peptide hormone that is secreted by atrial stretch and regulates blood pressure and volume by inhibiting reabsorption of sodium by the kidney (349). Immunochemical studies showed that ANF is present in acinar cells. Early studies suggested that ANF did not influence protein or fluid secretion. However, incubation of rat acini with ANF caused a dose-dependent elevation of cellular cGMP (117) showing that guanylate cyclase receptors transduce ANF signaling (210). Injection of human ANF in dogs induced bicarbonate but not sodium or protein secretion (262). ANF also stimulates pancreatic natriuretic peptide receptor-C (NPR-C)-mediated phosphoinositide-dependent pathway in rats, causing the release of fluid and protein (297). NPR-C is a non-guanylyl cyclase receptor and is coupled to adenylyl cyclase inhibition or phospholipase C activation through Gi proteins. ANF attenuated secretin- and VIP-induced elevation of intracellular cAMP and this effect was blocked by inhibitors of protein kinase C and phospholipase C (296). Along with elevating intracellular cAMP, secretin mediates the efflux of cAMP from intact pancreas and acinar cells. ANF augmented secretin-induced cAMP efflux and caused the rapid elimination of cAMP from cells. The multidrug resistance protein 4 (MRP4) has been reported to play a role in the extrusion of cAMP in many cellular systems. MRP4 is also expressed in the pancreas and genetic knockdown of MRP4 expression reduced intracellular cAMP levels in acinar cells by ANF and an NPR-C dependent mechanism (290).
C-natriuretic peptide is structurally similar to ANF and is expressed in the CNS and gastrointestinal tract. CNP increases pancreatic protein, chloride, and fluid secretion (without influencing bicarbonate output) suggesting that it acts on acinar rather than duct cells. Truncal vagotomy or perivagal application of capsaicin or hexamethonium attenuated chloride secretion, demonstrating that the effect of CNP is modulated by the parasympathetic nervous system (295). At low concentrations, CNP induced protein secretion by activation of NPR-C. Similar to ANF, CNP-induced amylase release was inhibited by PLC and PKC inhibitors. CNP also elevated intracellular cGMP and reduced cAMP concentrations suggesting that CNP can interact directly with receptors located on pancreatic acini (294).
Insulin
Insulin modulates pancreatic exocrine function and insulin receptors are present in high density on the basolateral surfaces of acinar cells (23). Insulin increases pancreatic enzyme synthesis and secretion and its effects are enhanced by CCK and secretin (4, 164, 189, 191, 216). Since CCK induces insulin release in the presence of glucose and amino acids, it is possible that these two hormones act in conjunction on exocrine stimulation following food intake (202, 298).
Limited data suggest that insulin regulates exocrine secretion by potentiation of ouabain-sensitive Na+,K+-ATPase and by vagal cholinergic input (113, 216, 280). The action of insulin on exocrine secretion is modulated by PP which exerts an inhibitory effect on pancreatic secretion (276). Since galanin, pancreastatin, and somatostatin are known to inhibit insulin secretion, it is possible that these peptides also regulate insulin-mediated amylase release (18, 190, 235).
Bombesin
Bombesin is a 14 amino acid peptide homolog of GRP and neuromedin B and has the ability to suppress appetite (355). The effects of bombesin on pancreatic exocrine secretion appear to vary based on the species. In pigs, administration of bombesin alone or in combination with secretin induced protein but not fluid secretion (195). In guinea pigs, bombesin was very effective in inducing bicarbonate release from interlobular ducts, and this effect was blocked by a GRP receptor antagonist (332). Administration of bombesin to rats resulted in pancreatic hypertrophy with increased pancreatic weight, protein, RNA, and enzyme content and this effect was not regulated by CCK or secretin (194, 330).
Melatonin
Melatonin is a lipophilic hormone produced by the pineal gland as well as by certain neuroendocrine cells located in the gastrointestinal tract. Melatonin receptors are present on acinar cells and melatonin protects the pancreas against caerulein-induced acute pancreatitis (148). Initial studies showed that melatonin induced pancreatic amylase release which was mediated by CCK, vagal sensory nerves, and melatonin type 2 receptors. However, melatonin did not appear to have a direct effect on pancreatic acinar cells (146, 147, 192, 256). The extent and importance of melatonin-induced pancreatic secretion is not well understood and has recently been proposed to involve activation of the entero-pancreatic reflex and CCK release from the duodenum (145).
Amylin
Amylin is a 37 amino acid hormone that is co-secreted along with insulin from pancreatic β-cells in response to nutrients. Amylin stimulates CCK-independent pancreatic secretion in rats, and this effect is blocked by proton pump inhibitors and atropine, perhaps due to inhibition of somatostatin release (85).Amylin was also shown to stimulate protein secretion from pancreatic AR42J cells by a mechanism involving activation of GPCRs and release of Ca2+ from intracellular stores (137). Others investigations have suggested that amylin has no effect on pancreatic exocrine secretion from isolated perfused pancreas, acinar preparations, or AR42J cells (77, 163, 367). Hence the effects of amylin on exocrine secretion remain unresolved.
Fibroblast Growth Factor 21 (FGF21)
Fibroblast Growth Factor 21 was initially discovered in the liver and thymus of mouse embryos (260), and was later found to be expressed at high levels in pancreatic acinar cells. FGF21 overexpression protected mice against caerulein and alcohol-induced pancreatitis (119, 153, 343). By evaluating the function of FGF21 in transgenic and knockout mice, it was discovered that FGF21 regulates pancreatic enzyme secretion from acinar cells in a paracrine/autocrine manner (52). FGF21 binds its heteromeric receptor (comprised of FGFR1c and β-klotho) to elevate intracellular calcium levels by signaling through phospholipase C and inositol trisphosphate. Zymogen granule accumulation was observed in both FGF21 knockout and β-klotho knockout mice. FGF21 is secreted in acinar cells in response to ER stress. Unlike CCK, FGF21’s role in acinar cells is limited to augmentation of amylase secretion, as it does not increase synthesis of pancreatic enzymes.
Histamine
The amino acid histamine is a potential mediator of pancreatic exocrine secretion, although it may have a gender-dependent role (322). Activation of H1 receptors and inhibition of H2 receptors in the rabbit pancreas led to an increase in fluid and protein secretion suggesting differential action based on regulation and coupling of the two receptors (275). The effect of histamine on pancreatic secretion is considered to be minor at best under normal physiological conditions.
C. Inhibitory Hormones
Peptide YY and Pancreatic Peptide
The NPY family of peptides consists of three hormones: NPY, PYY, and PP (95, 357). All three peptides contain 36 residues several of which are tyrosines and share a tertiary structural motif known as the PP fold. The N-terminal amino acids of PYY and NPY can be cleaved by peptidases to generate truncated forms, PYY3-36 and NPY3-36,which are biologically active. NPY has been localized to sympathetic pancreatic nerves and its role has been discussed previously in this review (42). In islets, PYY is coexpressed with glucagon in α cells, whereas PP is secreted postprandially by F cells of the islets of Langerhans. In certain species, PP immunopositive cells are also present in the exocrine pancreas and some of these PP cells also express PYY (72). These three hormones exert their effects through a family of five GPCRs denoted Y1-5. NPY and PYY possess similar affinity for Y1, Y2 and Y5, PYY3-36 interacts preferentially with Y2, whereas PP is the preferred ligand for Y4 (132).
PYY levels in blood are elevated postprandially and following instillation of fatty acids into the distal small intestine (214). Physiologically relevant concentrations of PYY in the circulation inhibit both meal- and hormone-stimulated pancreatic secretion (273, 274, 337). Intravenous administration of PYY significantly diminishes secretin- and secretin plus CCK-mediated pancreatic protein and fluid secretion concomitant with a reduction in pancreatic blood flow (140, 151, 274, 331). However, PYY does not inhibit 2-diacylglycerol stimulated pancreatic secretion, suggesting that suppression of CCK-stimulated exocrine secretion occurs prior to activation by 2-diacylglycerol or does not involve protein kinase C-activated signaling (67). In denervated pancreas, PYY1-36 but not PYY3-36,reduced CCK-stimulated amylase release suggesting that hormonal effects of PYY are mediated by Y1 receptors (62, 97). Autoradiographic analysis of rat pancreas with radio-iodinated PYY ligand demonstrated that Y1 receptors are present primarily on smooth muscle cells of blood vessels. Specific staining was not observed in acinar cells indicating that decreased protein and fluid secretion could be due to reduced blood flow (311).
PP secretion is also stimulated by ingesting a meal and can be reproduced by intraduodenal infusion of acid, aromatic amino acids, or fatty acids (22, 50, 161, 306, 338). Like PYY, PP attenuated secretin- and CCK-mediated exocrine secretion in dogs independent of cholinergic blockade (61, 62, 171). In one study, PP decreased secretin- and secretin plus CCK-mediated amylase release in dispersed acini, suggesting that PP can act directly on acinar cells (152). However, although bovine and rat PP inhibited CCK-stimulated protein secretion in vivo, both peptides were ineffective in vitro, and binding of bovine PP to rat acinar cells or lobules was not observed (208). Other studies have also not shown direct effects of PP (357). In humans, unlike dogs, infusion of PP decreased pancreatic protein output, but did not influence bicarbonate secretion suggesting species-specific differences in PP action (175). However, unlike PYY, PP does not affect pancreatic blood flow and therefore inhibits exocrine secretion at the cellular level (61, 62, 171).
Somatostatin
Somatostatin is composed of 14 or 28 amino acids and is produced by δ cells of pancreatic islets. It is also secreted by certain intestinal cells and by neurons in the hypothalamus. It is released into the blood after a meal but functions primarily through a paracrine mechanism. It has broad inhibitory actions on the release of several hormones and their target organs.
Somatostatin and its analogs inhibited secretin- and CCK-induced protein secretion in a dose-dependent fashion. Low doses of somatostatin exerted a greater inhibitory effect on CCK-stimulated pancreatic secretion compared to secretin-stimulated secretion (190, 203, 314, 315). Secretin-mediated activation of slowly-activating voltage-dependent K+ channels (present on the basolateral surface of pancreatic acini) resulted in cAMP generation and secretion of chloride ions. Addition of somatostatin to acini, decreased intracellular cAMP production as well as secretin-mediated enhancement of K+ current suggesting that somatostatin regulates exocrine secretion through this pathway (187). Additionally, somatostatin inhibited Ca2+-dependent and cAMP-stimulated amylase release by inhibiting exocytosis through a Gi protein-dependent mechanism (263).
Somatostatin also inhibits exocrine secretion via a neural mechanism. Based on atropine, hexamethonium, and tetrodotoxin sensitivities it appears that peptidergic but not cholinergic and nicotinic acetylcholine receptors present on sympathetic and parasympathetic ganglia mediate somatostatin action (248). Somatostatin-mediated inhibition of secretin-stimulated fluid and protein secretion was not influenced by denervation, suggesting that extrapancreatic nerves are not involved. Bethanechol, a muscarinic receptor agonist, reversed the inhibitory effects of somatostatin, indicating that its actions are mediated primarily by intrapancreatic cholinergic neurons (184). The mechanisms by which somatostatin inhibits pancreatic secretion are not completely understood. However, it is believed that somatostatin has an inhibitory effect on the release of hormones and neurotransmitters that normally stimulate pancreatic secretion.
Galanin
Galanin is a 29 amino acid peptide that plays diverse roles including inhibition of insulin, somatostatin, and PP secretion from the pancreas (16). It is found in the secretory granules of central and peripheral neurons suggesting that it functions as a neurotransmitter. Galanin immunoreactivity was present in nerve fibers surrounding acini, ductules, and blood vessels, with 73% of fibers being dual positive for galanin and VIP (313). Galanin receptor 3 mRNA is present in acinar cells indicating that galanin may act directly on acini (17). Consistent with this finding, galanin inhibited CCK- and carbachol-stimulated amylase release from acinar cells (5). Galanin inhibited the sustained phase of amylase release stimulated by carbachol, suggesting that it attenuates cholinergic action possibly by a mechanism that involves pertussis toxin-sensitive Gi proteins (18, 82, 120, 157). Extrapancreatic nerves are not involved in its action since galanin inhibited food-, secretin- and CCK-mediated fluid release, as well as food- and CCK-mediated protein release in both innervated and denervated dogs (35).
Pancreastatin
Pancreastatin is derived from the cleavage of chromogranin A and is expressed in many neuroendocrine tissues. It has been localized to duct cells of the exocrine pancreas and its numerous roles include inhibition of pancreatic exocrine secretion (1). Initial studies showed that pancreastatin inhibited postprandial fluid and protein secretion in rats with bile-pancreatic juice diversion. No effect was observed on basal secretion, secretin-stimulated secretion in conscious rats, or CCK-stimulated secretion from dispersed acini. However, pancreastatin inhibited CCK-stimulated pancreatic secretion in conscious rats although it did not influence plasma CCK levels. These results suggest that pancreastatin does not have a direct effect on acinar cells, but may regulate the intestinal phase of pancreatic secretion (86, 233, 235, 350). Pancreastatin inhibited caerulein-induced blood flow in the exocrine pancreas raising the possibility that its inhibitory effects are derived from its role in regulating pancreatic blood flow (231).
Glucagon
Glucagon is released from the endocrine pancreas after ingestion of a meal ((33).Early investigations suggested that glucagon inhibited secretin- or secretin- and CCK-stimulated pancreatic protein but not bicarbonate secretion (51, 232). However other studies have demonstrated that glucagon inhibits postprandial protein and bicarbonate secretion (83, 111, 321). The effect of glucagon on isolated pancreatic lobules and acini appears to be direct and stimulatory, instead of inhibitory, suggesting complex action at the cellular versus physiological levels (2, 133, 271, 324). The experimentally observed effects of glucagon on exocrine secretion are inconclusive and merit further investigation. Another proglucagon derived peptide known as oxyntomodulin, is a dual agonist of GLP-1R and glucagon receptors. Although this peptide did not influence plasma amylase concentration on its own, it attenuated CCK-mediated plasma amylase release. The mechanism of action of oxyntomodulin has not been identified (6).
Ghrelin
Ghrelin is a 28 amino acid orexigenic hormone released by gastric endocrine cells under fasting conditions. Ghrelin stimulates acid secretion by oxyntic cells in the stomach, and plasma levels of ghrelin rise immediately before a meal suggesting a role in modulating ingestive behavior. In the pancreas both ghrelin and its receptor have been identified in acini by evaluation of protein and mRNA expression (185). Ghrelin expression was not altered by gastric acid inhibition, acute pancreatitis, or food deprivation although its receptor was upregulated by gastric acid inhibition and downregulated during acute pancreatitis (185). Experimentally, ghrelin did not affect basal or CCK-stimulated amylase release from dispersed acini. However, ghrelin inhibited CCK-stimulated protein secretion in normal and vagotomized rats and amylase secretion from lobules exposed to depolarizing potassium concentrations, suggesting that it modulates intrapancreatic neurons (368).
Leptin
Leptin is a 16 kDa orexigenic peptide that is secreted by adipocytes and regulates energy homeostasis by reducing food intake while increasing energy expenditure. In the pancreas, intravenous or intraperitoneal administration of leptin reduced basal and CCK-stimulated protein output in vivo. This effect was attenuated by CCK-1 receptor blockade, vagotomy, and capsaicin pretreatment suggesting that it inhibited pancreatic exocrine secretion through a CCK-dependent vagal pathway. Leptin had no effect on dispersed acini in vitro, further supporting a neural mechanism of action (144, 217). In contrast, intraduodenal infusion of leptin in fasted rats augmented pancreatic protein output possibly by elevating plasma CCK levels leading to activation of sensory neurons (254).
Adrenomedullin
Adrenomedullin is colocalized with PP in F cells of pancreatic islets and inhibits insulin (207). In one study, it interacted directly with acinar cells and inhibited CCK-stimulated pancreatic amylase release (342). The mechanism of adrenomedullin action is not well understood.
IV. CONCLUSION
Pancreatic secretion is a complex process that is initiated by the sight and smell of food and progresses until food enters the duodenum. At each level of food digestion, this process is regulated by various stimuli which affect neuronal and hormonal pathways. These pathways are both stimulatory and inhibitory and optimize the release of enzymes, bicarbonate, and fluid.
V. REFERENCES
- Adeghate E, Ember Z, Donath T, Pallot DJ, and Singh J. Immunohistochemical identification and effects of atrial natriuretic peptide, pancreastatin, leucine-enkephalin, and galanin in the porcine pancreas. Peptides 17: 503-509, 1996. PMID: 8735979.
- Adler G. Effect of glucagon on the secretory process in the rat exocrine pancreas. Cell Tissue Res 182: 193-204, 1977. PMID: 902302.
- Adler G, Beglinger C, Braun U, Reinshagen M, Koop I, Schafmayer A, Rovati L, and Arnold R. Interaction of the cholinergic system and cholecystokinin in the regulation of endogenous and exogenous stimulation of pancreatic secretion in humans. Gastroenterology 100: 537-543, 1991. PMID: 1702077.
- Adler G and Kern HF. Regulation of exocrine pancreatic secretory process by insulin in vivo. Horm Metab Res 7: 290-296, 1975. PMID: 807510.
- Ahren B, Andren-Sandberg A, and Nilsson A. Galanin inhibits amylase secretion from isolated rat pancreatic acini. Pancreas 3: 559-562, 1988. PMID: 2460854.
- Akalestou E, Christakis I, Solomou AM, Minnion JS, Rutter GA, and Bloom SR. Proglucagon-Derived Peptides Do Not Significantly Affect Acute Exocrine Pancreas in Rats. Pancreas 45: 967-973, 2016. PMID: 26731187.
- Alcon S, Rosado JA, Garcia LJ, Pariente JA, Salido GM, and Pozo MJ. Secretin potentiates guinea pig pancreatic response to cholecystokinin by a cholinergic mechanism. Can J Physiol Pharmacol 74: 1342-1350, 1996. PMID: 9047045.
- Alonso RM, Rodriguez AM, Garcia LJ, Lopez MA, and Calvo JJ. Comparison between the effects of VIP and the novel peptide PACAP on the exocrine pancreatic secretion of the rat. Pancreas 9: 123-128, 1994. PMID: 7509062.
- Anagnostides A, Chadwick VS, Selden AC, and Maton PN. Sham feeding and pancreatic secretion. Evidence for direct vagal stimulation of enzyme output. Gastroenterology 87: 109-114, 1984. PMID: 6724252.
- Ashton N, Argent BE, and Green R. Effect of vasoactive intestinal peptide, bombesin and substance P on fluid secretion by isolated rat pancreatic ducts. J Physiol 427: 471-482, 1990. PMID: 1698981.
- Babic T, Browning KN, Kawaguchi Y, Tang X, and Travagli RA. Pancreatic insulin and exocrine secretion are under the modulatory control of distinct subpopulations of vagal motoneurones in the rat. J Physiol 590: 3611-3622, 2012. PMID: 22711959.
- Babic T, and Travagli RA. Neural Control of the Pancreas. Pancreapedia: Exocrine Pancreas Knowledge Base, 2016. DOI: 10.3998/panc.2016.27.
- Baca I, Feurle GE, Haas M, and Mernitz T. Interaction of neurotensin, cholecystokinin, and secretin in the stimulation of the exocrine pancreas in the dog. Gastroenterology 84: 556-561, 1983. PMID: 6295872.
- Barlow TE, Greenwell JR, Harper AA, and Scratcherd T. The effect of adrenaline and noradrenaline on the blood flow, electrical conductance and external secretion of the pancreas. J Physiol 217: 665-678, 1971. PMID: 4398605.
- Barnhart DC, Sarosi GA, Jr., and Mulholland MW. PACAP-38 causes phospholipase C-dependent calcium signaling in rat acinar cell line. Surgery 122: 465-474, 1997. PMID: 9288154.
- Barreto SG. Galanin. In: The Pancreapedia: Exocrine Pancreas Knowledge Base, 2015. DOI: 10.3998/panc.2015.21.
- Barreto SG, Bazargan M, Zotti M, Hussey DJ, Sukocheva OA, Peiris H, Leong M, Keating DJ, Schloithe AC, Carati CJ, Smith C, Toouli J, and Saccone GT. Galanin receptor 3--a potential target for acute pancreatitis therapy. Neurogastroenterol Motil 23: e141-151, 2011. PMID: 21303427.
- Barreto SG, Woods CM, Carati CJ, Schloithe AC, Jaya SR, Toouli J, and Saccone GT. Galanin inhibits caerulein-stimulated pancreatic amylase secretion via cholinergic nerves and insulin. Am J Physiol Gastrointest Liver Physiol 297: G333-339, 2009. PMID: 19497960.
- Bastie MJ, Vaysse N, Brenac B, Pascal JP, and Ribet A. Effects of catecholamines and their inhibitors on the isolated canine pancreas. II. Dopamine. Gastroenterology 72: 719-723, 1977. PMID: 838228.
- Becker S, Niebel W, and Singer MV. Nervous control of gastric and pancreatic secretory response to 2-deoxy-D-glucose in the dog. Digestion 39: 187-196, 1988. PMID: 3209002.
- Beglinger C, Fried M, Whitehouse I, Jansen JB, Lamers CB, and Gyr K. Pancreatic enzyme response to a liquid meal and to hormonal stimulation. Correlation with plasma secretin and cholecystokinin levels. J Clin Invest 75: 1471-1476, 1985. PMID: 3998145.
- Beglinger C, Taylor IL, Grossman MI, and Solomon TE. Pancreatic polypeptide release: role of stimulants of exocrine pancreatic secretion in dogs. Gastroenterology 87: 530-536, 1984. PMID: 6146550.
- Bergeron JJ, Rachubinski R, Searle N, Sikstrom R, Borts D, Bastian P, and Posner BI. Radioautographic visualization of in vivo insulin binding to the exocrine pancreas. Endocrinology 107: 1069-1080, 1980. PMID: 6997018.
- Bilski J, Konturek PK, Krzyzek E, and Konturek SJ. Feedback control of pancreatic secretion in rats. Role of gastric acid secretion. J Physiol Pharmacol 43: 237-257, 1992. PMID: 1493255.
- Bissonnette BM, Collen MJ, Adachi H, Jensen RT, and Gardner JD. Receptors for vasoactive intestinal peptide and secretin on rat pancreatic acini. Am J Physiol 246: G710-717, 1984. PMID: 6204536.
- Bjornsson OG, Maton PN, Fletcher DR, and Chadwick VS. Effects of duodenal perfusion with sodium taurocholate on biliary and pancreatic secretion in man. Eur J Clin Invest 12: 97-105, 1982. PMID: 6807694.
- Blair EL, Brown JC, Harper AA, and Scratcherd T. A gastric phase of pancreatic secretion. J Physiol 184: 812-824, 1966. PMID: 5912208.
- Blomfield J and Settree PJ. Ultrastructural responses of rat exocrine pancreas to cholecystokinin octapeptide and secretin. Exp Mol Pathol 38: 389-397, 1983. PMID: 6303836.
- Bohorquez DV, Samsa LA, Roholt A, Medicetty S, Chandra R, and Liddle RA. An enteroendocrine cell-enteric glia connection revealed by 3D electron microscopy. PLoS One 9: e89881, 2014. PMID: 24587096.
- Bohorquez DV, Shahid RA, Erdmann A, Kreger AM, Wang Y, Calakos N, Wang F, and Liddle RA. Neuroepithelial circuit formed by innervation of sensory enteroendocrine cells. J Clin Invest 125: 782-786, 2015. PMID: 25555217.
- Bondesen S, Christensen H, Lindorff-Larsen K, and Schaffalitzky de Muckadell OB. Plasma secretin in response to pure bile salts and endogenous bile in man. Dig Dis Sci 30: 440-444, 1985. PMID: 3987477.
- Bouras EP, Misukonis MA, and Liddle RA. Role of calcium in monitor peptide-stimulated cholecystokinin release from perifused intestinal cells. Am J Physiol 262: G791-796, 1992. PMID: 1590389.
- Bozadjieva N, Williams JA, and Bernal-Mizrachi E. Glucagon. In: The Pancreapedia: Exocrine Pancreas Knowledge Base, 2013. DOI: 10.3998/panc.2013.23.
- Bozkurt T, Adler G, Koop I, and Arnold R. Effect of atropine on intestinal phase of pancreatic secretion in man. Digestion 41: 108-115, 1988. PMID: 2464512.
- Brodish RJ, Kuvshinoff BW, Fink AS, and McFadden DW. Inhibition of pancreatic exocrine secretion by galanin. Pancreas 9: 297-303, 1994. PMID: 7517543.
- p class="EndNoteBibliography">36. Brooks AM and Grossman MI. Postprandial pH and neutralizing capacity of the proximal duodenum in dogs. Gastroenterology 59: 85-89, 1970. PMID: 5426993.
- Bunnett NW, Mulvihill SJ, and Debas HT. Calcitonin gene-related peptide inhibits exocrine secretion from the rat pancreas by a neurally mediated mechanism. Exp Physiol 76: 115-123, 1991. PMID: 2015068.
- Burhol PG, Lygren I, Waldum HL, and Jorde R. The effect of duodenal infusion of bile on plasma VIP, GIP, and secretin and on duodenal bicarbonate secretion. Scand J Gastroenterol 15: 1007-1011, 1980. PMID: 7233065.
- Burnham DB, McChesney DJ, Thurston KC, and Williams JA. Interaction of cholecystokinin and vasoactive intestinal polypeptide on function of mouse pancreatic acini in vitro. J Physiol 349: 475-482, 1984. PMID: 6204039.
- Camello PJ and Salido GM. Inhibitory interactions between stimulus-secretion pathways in the exocrine rat pancreas. Biochem Pharmacol 46: 1005-1009, 1993. PMID: 8216342.
- Cargill JM and Wormsley KG. Effect of gastric distension on human pancreatic secretion. Acta Hepatogastroenterol (Stuttg) 26: 235-238, 1979. PMID: 484173.
- Carlei F, Allen JM, Bishop AE, Bloom SR, and Polak JM. Occurrence, distribution and nature of neuropeptide Y in the rat pancreas. Experientia 41: 1554-1557, 1985. PMID: 2866975.
- Chandra R, Samsa L, Vigna S, and Liddle R. Pseudopod-like basal cell processes in intestinal cholecystokinin cells. Cell Tiss Res 341: 289-297, 2010. PMID: 20582553.
- Chandra R, Wang Y, Shahid RA, Vigna SR, Freedman NJ, and Liddle RA. Immunoglobulin-like domain containing receptor 1 mediates fat-stimulated cholecystokinin secretion. J Clin Invest 123: 3343-3352, 2013. PMID: 23863714.
- Chariot J, Appia F, del Tacca M, Tsocas A, and Roze C. Central and peripheral inhibition of exocrine pancreatic secretion by alpha-2 adrenergic agonists in the rat. Pharmacol Res Commun 20: 707-717, 1988. PMID: 2905481.
- Chariot J, Roze C, de la Tour J, Souchard M, and Vaille C. Modulation of stimulated pancreatic secretion by sympathomimetic amines in the rat. Pharmacology 26: 313-323, 1983. PMID: 6878424.
- Chayvialle JA, Miyata M, Rayford PL, and Thompson JC. Effects of test meal, intragastric nutrients, and intraduodenal bile on plasma concentrations of immunoreactive somatostatin and vasoactive intestinal peptide in dogs. Gastroenterology 79: 844-852, 1980. PMID: 6106620.
- Chey WY and Chang T-M. Secretin, 100 years later. J Gastroenterol 38: 1025-1035, 2003. PMID: 14673718.
- Chey WY and Chang T. Neural hormonal regulation of exocrine pancreatic secretion. Pancreatology 1: 320-335, 2001. PMID: 12120211.
- Choi BR and Palmquist DL. High fat diets increase plasma cholecystokinin and pancreatic polypeptide, and decrease plasma insulin and feed intake in lactating cows. J Nutr 126: 2913-2919, 1996. PMID: 8914965.
- Clain JE, Barbezat GO, Waterworth MM, and Bank S. Glucagon inhibition of secretin and combined secretin and cholecystokinin stimulated pancreatic exocrine secretion in health and disease. Digestion 17: 11-17, 1978. PMID: 627317.
- Coate KC, Hernandez G, Thorne CA, Sun S, Le TDV, Vale K, Kliewer SA, and Mangelsdorf DJ. FGF21 Is an Exocrine Pancreas Secretagogue. Cell Metabolism 25: 472-480, 2017. PMID: 28089565.
- Cooke AR, Nahrwold DL, and Grossman MI. Diversion of pancreatic juice on gastric and pancreatic response to a meal stimulus. Am J Physiol 213: 637-639, 1967. PMID: 6036779.
- Criddle DN, Booth DM, Mukherjee R, McLaughlin E, Green GM, Sutton R, Petersen OH, and Reeve JR. Cholecystokinin-58 and cholecystokinin-8 exhibit similar actions on calcium signaling, zymogen secretion, and cell fate in murine pancreatic acinar cells. Am J Physiol Gastrointest Liver Physiol 297: G1085-G1092, 2009. PMID: 19815626.
- Dale WE, Turkelson CM, and Solomon TE. Role of cholecystokinin in intestinal phase and meal-induced pancreatic secretion. Am J Physiol 257: G782-790, 1989. PMID: 2480719.
- De Giorgio R, Sternini C, Brecha NC, Widdison AL, Karanjia ND, Reber HA, and Go VL. Patterns of innervation of vasoactive intestinal polypeptide, neuropeptide Y, and gastrin-releasing peptide immunoreactive nerves in the feline pancreas. Pancreas 7: 376-384, 1992. PMID: 1594560.
- Debas HT and Grossman MI. Pure cholecystokinin: pancreatic protein and bicarbonate response. Digestion 9: 469-481, 1973. PMID: 4787845.
- Debas HT and Yamagishi T. Evidence for pyloropancreatic reflux for pancreatic exocrine secretion. Am J Physiol 234: E468-471, 1978. PMID: 645898.
- Defilippi C, Solomon TE, and Valenzuela JE. Pancreatic secretory response to sham feeding in humans. Digestion 23: 217-223, 1982. PMID: 6183160.
- Delcenserie R, Devaux MA, and Sarles H. Action of dopamine on the exocrine pancreatic secretion of the intact dog. Br J Pharmacol 88: 189-195, 1986. PMID: 2871881.
- DeMar AR, Lake R, and Fink AS. The effect of pancreatic polypeptide and peptide YY on pancreatic blood flow and pancreatic exocrine secretion in the anesthetized dog. Pancreas 6: 9-14, 1991. PMID: 1994383.
- DeMar AR, Taylor IL, and Fink AS. Pancreatic polypeptide and peptide YY inhibit the denervated canine pancreas. Pancreas 6: 419-426, 1991. PMID: 1876598.
- Dembinski A, Konturek SJ, and Thor P. Gastric and pancreatic responses to meals varying in pH. J Physiol 243: 115-128, 1974. PMID: 4449058.
- Demol P and Sarles H. Action of catecholamines on exocrine pancreatic secretion of conscious rats. Arch Int Pharmacodyn Ther 243: 149-163, 1980. PMID: 7387257.
- Demol P and Sarles H. Action of fatty acids on the exocrine pancreatic secretion of the conscious rat: further evidence for a protein pancreatic inhibitory factor. J Physiol 275: 27-37, 1978. PMID: 633115.
- Demol P and Sarles H. Inhibition of exocrine pancreatic secretion by alpha-adrenergic blocking agents in conscious rats. Arch Int Pharmacodyn Ther 243: 164-176, 1980. PMID: 6104476.
- Deng X, Guarita DR, Wood PG, Kriess C, and Whitcomb DC. PYY potently inhibits pancreatic exocrine secretion mediated through CCK-secretin-stimulated pathways but not 2-DG-stimulated pathways in awake rats. Dig Dis Sci 46: 156-165, 2001. PMID: 11270780.
- DiMagno EP, Go VL, and Summerskill HJ. Intraluminal and postabsorptive effects of amino acids on pancreatic enzyme secretion. J Lab Clin Med 82: 241-248, 1973. PMID: 4721379.
- Doi R, Inoue K, Kogire M, Sumi S, Yun M, Futaki S, Fujii N, Yajima H, and Tobe T. Effect of synthetic human cholecystokinin-33 on exocrine pancreas. Biochem Biophys Res Commun 150: 1251-1255, 1988. PMID: 3342068.
- Eberlein GA, Eysselein VE, Hesse WH, Goebell H, Schaefer M, and Reeve JR, Jr. Detection of cholecystokinin-58 in human blood by inhibition of degradation. Am J Physiol 253: G477-482, 1987. PMID: 3661709.
- Ekblad E, Alm P, and Sundler F. Distribution, origin and projections of nitric oxide synthase-containing neurons in gut and pancreas. Neuroscience 63: 233-248, 1994. PMID: 7534882.
- Ekblad E and Sundler F. Distribution of pancreatic polypeptide and peptide YY. Peptides 23: 251-261, 2002. PMID: 11825640.
- Ember Z, Yago MD, and Singh J. Distribution of nitric oxide synthase and secretory role of exogenous nitric oxide in the isolated rat pancreas. Int J Pancreatol 29: 77-84, 2001. PMID: 11876252.
- Eysselein VE, Eberlein GA, Hesse WH, Singer MV, Goebell H, and Reeve JR, Jr. Cholecystokinin-58 is the major circulating form of cholecystokinin in canine blood. J Biol Chem 262: 214-217, 1987. PMID: 3793725.
- Fahrenkrug J, Schaffalitzky de Muckadell OB, Holst JJ, and Jensen SL. Vasoactive intestinal polypeptide in vagally mediated pancreatic secretion of fluid and HCO3. Am J Physiol 237: E535-540, 1979. PMID: 517650.
- Faichney A, Chey WY, Kim YC, Lee KY, Kim MS, and Chang TM. Effect of sodium oleate on plasma secretin concentration and pancreatic secretion in dog. Gastroenterology 81: 458-462, 1981. PMID: 7250635.
- Fehmann H-C, Weber V, Göke R, Göke B, Eissele R, and Arnold R. Islet amyloid polypeptide (IAPP; Amylin) influences the endocrine but not the exocrine rat pancreas. Biochem Biophys Res Comm 167: 1102-1108, 1990. PMID: 1690993.
- Feurle GE and Reinecke M. Neurotensin interacts with carbachol, secretin, and caerulein in the stimulation of the exocrine pancreas of the rat in vitro. Regul Pept 7: 137-143, 1983. PMID: 6197722.
- Fink AS, Luxenburg M, and Meyer JH. Regionally perfused fatty acids augment acid-induced canine pancreatic secretion. Am J Physiol 245: G78-84, 1983. PMID: 6869548.
- Fink AS and Meyer JH. Intraduodenal emulsions of oleic acid augment acid-induced canine pancreatic secretion. Am J Physiol 245: G85-91, 1983. PMID: 6869549.
- Fink AS, Miller JC, Jehn DW, and Meyer JH. Digests of protein augment acid-induced canine pancreatic secretion. Am J Physiol 242: G634-641, 1982. PMID: 6807099.
- Flowe KM, Lally KM, and Mulholland MW. Galanin inhibits rat pancreatic amylase release via cholinergic suppression. Peptides 13: 487-492, 1992. PMID: 1381828.
- Fontana G, Costa PL, Tessari R, and Labo G. Effect of glucagon on pure human exocrine pancretic secretion. Am J Gastroenterol 63: 490-494, 1975. PMID: 1146806.
- Fridolf T, Sundler F, and Ahren B. Pituitary adenylate cyclase-activating polypeptide (PACAP): occurrence in rodent pancreas and effects on insulin and glucagon secretion in the mouse. Cell Tissue Res 269: 275-279, 1992. PMID: 1423494.
- Funakoshi A, Miyasaka K, Kitani K, Nakamura J, Funakoshi S, Fukuda H, and Fujii N. Stimulatory effects of islet amyloid polypeptide (amylin) on exocrine pancreas and gastrin release in conscious rats. Regul Pept 38: 135-143, 1992. PMID: 1574608.
- Funakoshi A, Miyasaka K, Nakamura R, Kitani K, and Tatemoto K. Inhibitory effect of pancreastatin on pancreatic exocrine secretion in the conscious rat. Regul Pept 25: 157-166, 1989. PMID: 2474177.
- Furukawa N and Okada H. Effects of antral distension on pancreatic exocrine secretion in dogs: evidence for a short reflex. Jpn J Physiol 37: 671-685, 1987. PMID: 3430872.
- Furukawa N and Okada H. Effects of stimulation of the hypothalamic area on pancreatic exocrine secretion in dogs. Gastroenterology 97: 1534-1543, 1989. PMID: 2684723.
- Furuta Y, Hashimoto K, and Washizaki M. beta-Adrenoceptor stimulation of exocrine secretion from the rat pancreas. Br J Pharmacol 62: 25-29, 1978. PMID: 202364.
- Gabryelewicz A, Kulesza E, and Konturek SJ. Comparison of loxiglumide, a cholecystokinin receptor antagonist, and atropine on hormonal and meal-stimulated pancreatic secretion in man. Scand J Gastroenterol 25: 731-738, 1990. PMID: 2396088.
- Gardner JD, Knight M, Sutliff VE, Tamminga CA, and Jensen RT. Derivatives of CCK-(26-32) as cholecystokinin receptor antagonists in guinea pig pancreatic acini. Am J Physiol 246: G292-295, 1984. PMID: 6199984.
- Gautam D, Han SJ, Heard TS, Cui Y, Miller G, Bloodworth L, and Wess J. Cholinergic stimulation of amylase secretion from pancreatic acinar cells studied with muscarinic acetylcholine receptor mutant mice. J Pharmacol Exp Ther 313: 995-1002, 2005. PMID: 15764735.
- Go VL, Hofmann AF, and Summerskill WH. Pancreozymin bioassay in man based on pancreatic enzyme secretion: potency of specific amino acids and other digestive products. J Clin Invest 49: 1558-1564, 1970. PMID: 5431665.
- Gomez GA, Upp JR, Jr., Lluis F, Alexander RW, Poston GJ, Greeley GH, Jr., and Thompson JC. Regulation of the release of cholecystokinin by bile salts in dogs and humans. Gastroenterology 94: 1036-1046, 1988. PMID: 3345873.
- Gomez GA and Greeley GHJ. Peptide YY. In: The Pancreapedia: Exocrine Pancreas Knowledge Base, 2013. DOI: 10.3998/panc.2013.6.
- Goyal D and Pisegna JR. Pituitary Adenylate Cyclase Activating Polypeptide (PACAP). Pancreapedia: Exocrine Pancreas Knowledge Base, 2014. DOI: 10.3998/panc.2014.12.
- Grandt D, Siewert J, Sieburg B, al Tai O, Schimiczek M, Goebell H, Layer P, Eysselein VE, Reeve JR, Jr., and Muller MK. Peptide YY inhibits exocrine pancreatic secretion in isolated perfused rat pancreas by Y1 receptors. Pancreas 10: 180-186, 1995. PMID: 7536329.
- Green G and Nasset ES. Effect of bile duct obstruction on pancreatic enzyme secretion and intestinal proteolytic enzyme activity in the rat. Am J Dig Dis 22: 437-444, 1977. PMID: 857664.
- Green GM. Role of gastric juice in feedback regulation of rat pancreatic secretion by luminal proteases. Pancreas 5: 445-451, 1990. PMID: 2199966.
- Green GM and Lyman RL. Feedback regulation of pancreatic enzyme secretion as a mechanism for trypsin inhibitor-induced hypersecretion in rats. Proc Soc Exp Biol Med 140: 6-12, 1972. PMID: 5033119.
- Green GM and Nasset ES. Role of dietary protein in rat pancreatic enzyme secretory response to a meal. J Nutr 113: 2245-2252, 1983. PMID: 6685174.
- Green GM, Taguchi S, Friestman J, Chey WY, and Liddle RA. Plasma secretin, CCK, and pancreatic secretion in response to dietary fat in the rat. Am J Physiol 256: G1016-1021, 1989. PMID: 2735407.
- Gries E, Hotz J, and Goebell H. Pancreatic exocrine secretion in response to intraduodenal infusion of different detergent agents in anesthetized cats. Digestion 34: 61-67, 1986. PMID: 3732639.
- Griesbacher T, Rainer I, Heinemann A, and Groisman D. Haemodynamic and exocrine effects of caerulein at submaximal and supramaximal levels on the rat pancreas: role of cholecystokinin receptor subtypes. Pancreatology 6: 65-75, 2006. PMID: 16327284.
- Guan D and Green GM. Significance of peptic digestion in rat pancreatic secretory response to dietary protein. Am J Physiol 271: G42-47, 1996. PMID: 8760105.
- Guan D, Spannagel A, Ohta H, Nakano I, Chey WY, and Green GM. Role of secretin in basal and fat-stimulated pancreatic secretion in conscious rats. Endocrinology 128: 979-982, 1991. PMID: 1989876.
- Hacki WH, Bloom SR, Mitznegg P, Domschke W, Domschke S, Belohlavek D, Demling L, and Wunsch E. Plasma secretin and pancreatic bicarbonate response to exogenous secretin in man. Gut 18: 191-195, 1977. PMID: 852751.
- Hajri A, Koenig M, Balboni G, and Damge C. Expression and characterization of gastrin-releasing peptide receptor in normal and cancerous pancreas. Pancreas 12: 25-35, 1996. PMID: 8927617.
- Hanssen LE, Hotz J, Hartmann W, Nehls W, and Goebell H. Immunoreactive secretin release following taurocholate perfusions of the cat duodenum. Scand J Gastroenterol 15: 89-95, 1980. PMID: 7367827.
- Hanssen LE, Hotz J, Layer P, and Goebell H. Bile-stimulated secretin release in cats. Scand J Gastroenterol 21: 886-890, 1986. PMID: 3775254.
- Harada H, Kochi F, Hanafusa E, Kobayashi T, Oka H, and Kimura I. Studies on the effect of glucagon on human pancreatic secretion by analysis of endoscopically obtained pure pancreatic juice. Gastroenterol Jpn 20: 28-36, 1985. PMID: 4018495.
- Hartmann W, Hotz J, Ormai S, Aufgebauer J, Schneider F, and Goebell H. Stimulation of bile and pancreatic secretion by duodenal perfusion with Na-taurocholate in the cat compared with jejunal and ileal perfusion. Scand J Gastroenterol 15: 433-442, 1980. PMID: 7433905.
- Hasegawa H, Okabayashi Y, Koide M, Kido Y, Okutani T, Matsushita K, Otsuki M, and Kasuga M. Effect of islet hormones on secretin-stimulated exocrine secretion in isolated perfused rat pancreas. Dig Dis Sci 38: 1278-1283, 1993. PMID: 8325188.
- Hashimoto K, Oguro K, and Furuta Y. Species difference in the secretory response to dopamine in the pancreas of dogs, cats, rabbits and rats. Arch Histol Jpn 40 Suppl: 129-132, 1977. PMID: 209761.
- Hegyi P, Maleth J, Venglovecz V, and Rakonczay Z, Jr. Pancreatic ductal bicarbonate secretion: challenge of the acinar Acid load. Front Physiol 2: 36, 2011. PMID: 21808623.
- Hegyi P, Rakonczay Z, Jr., Tiszlavicz L, Varro A, Toth A, Racz G, Varga G, Gray MA, and Argent BE. Protein kinase C mediates the inhibitory effect of substance P on HCO3- secretion from guinea pig pancreatic ducts. Am J Physiol Cell Physiol 288: C1030-1041, 2005. PMID: 15625303.
- Heisler S, Kopelman H, Chabot JG, and Morel G. Atrial natriuretic factor and exocrine pancreas: effects on the secretory process. Pancreas 2: 243-251, 1987. PMID: 2442744.
- Henriksen FW and Worning H. External pancreatic response to food and its relation to the maximal secretory capacity in dogs. Gut 10: 209-214, 1969. PMID: 5781141.
- Hernandez G, Luo T, Javed TA, Wen L, Kalwat MA, Vale K, Ammouri F, Husain SZ, Kliewer SA, and Mangelsdorf DJ. Pancreatitis is an FGF21-deficient state that is corrected by replacement therapy. Sci Transl Med 12: eaay5186, 2020. PMID: 31915301.
- Herzig KH, Brunke G, Schon I, Schaffer M, and Folsch UR. Mechanism of galanin's inhibitory action on pancreatic enzyme secretion: modulation of cholinergic transmission--studies in vivo and in vitro. Gut 34: 1616-1621, 1993. PMID: 7694889.
- Herzig KH, Schon I, Tatemoto K, Ohe Y, Li Y, Folsch UR, and Owyang C. Diazepam binding inhibitor is a potent cholecystokinin-releasing peptide in the intestine. Proc Natl Acad Sci USA 93: 7927-7932, 1996. PMID: 8755579.
- Hildebrand P, Beglinger C, Gyr K, Jansen JB, Rovati LC, Zuercher M, Lamers CB, Setnikar I, and Stalder GA. Effects of a cholecystokinin receptor antagonist on intestinal phase of pancreatic and biliary responses in man. J Clin Invest 85: 640-646, 1990. PMID: 2312719.
- Hira T, Nakajima S, Eto Y, and Hara H. Calcium-sensing receptor mediates phenylalanine-induced cholecystokinin secretion in enteroendocrine STC-1 cells. FEBS J 275: 4620-4626, 2008. PMID: 18691347.
- Hiramatsu K and Oshima K. Immunocytochemical study on the innervation of the chicken pancreas by vasoactive intestinal polypeptide (VIP)-containing nerves. Histol Histopathol 12: 961-965, 1997. PMID: 9302555.
- Hiraoka T, Fukuwatari T, Imaizumi M, and Fushiki T. Effects of oral stimulation with fats on the cephalic phase of pancreatic enzyme secretion in esophagostomized rats. Physiol Behav 79: 713-717, 2003. PMID: 12954413.
- Holst JJ, Fahrenkrug J, Knuhtsen S, Jensen SL, Poulsen SS, and Nielsen OV. Vasoactive intestinal polypeptide (VIP) in the pig pancreas: role of VIPergic nerves in control of fluid and bicarbonate secretion. Regul Pept 8: 245-259, 1984. PMID: 6379759.
- Holst JJ, Knuhtsen S, Jensen SL, Fahrenkrug J, Larsson LI, and Nielsen OV. Interrelation of nerves and hormones in stomach and pancreas. Scand J Gastroenterol Suppl 82: 85-99, 1983. PMID: 6138853.
- Holst JJ, Orskov C, Knuhtsen S, Sheikh S, and Nielsen OV. On the regulatory functions of neuropeptide Y (NPY) with respect to vascular resistance and exocrine and endocrine secretion in the pig pancreas. Acta Physiol Scand 136: 519-526, 1989. PMID: 2675536.
- Holst JJ, Rasmussen TN, Harling H, and Schmidt P. Effect of intestinal inhibitory peptides on vagally induced secretion from isolated perfused porcine pancreas. Pancreas 8: 80-87, 1993. PMID: 7678328.
- Holst JJ, Rasmussen TN, and Schmidt P. Role of nitric oxide in neurally induced pancreatic exocrine secretion in pigs. Am J Physiol 266: G206-213, 1994. PMID: 8141293.
- Holtmann G, Singer MV, Kriebel R, Stacker KH, and Goebell H. Differential effects of acute mental stress on interdigestive secretion of gastric acid, pancreatic enzymes, and gastroduodenal motility. Dig Dis Sci 34: 1701-1707, 1989. PMID: 2582983.
- Holzer P, Reichmann F, and Farzi A. Neuropeptide Y, peptide YY and pancreatic polypeptide in the gut-brain axis. Neuropeptides 46: 261-274, 2012. PMID: 22979996.
- Horiuchi A, Iwatsuki K, Ren LM, Kuroda T, and Chiba S. Dual actions of glucagon: direct stimulation and indirect inhibition of dog pancreatic secretion. Eur J Pharmacol 237: 23-30, 1993. PMID: 7689468.
- Hosotani R, Inoue K, Fujii N, Yajima H, and Tobe T. Effect of synthetic neuromedin C, a decapeptide of gastrin-releasing peptide (GRP [18-27]), on blood flow and exocrine secretion of the pancreas in dogs. Life Sci 36: 2429-2434, 1985. PMID: 3892215.
- Hosotani R, Inoue K, Kogire M, Suzuki T, Otsuki M, Rayford PL, Yajima H, and Tobe T. Synthetic neuromedin C stimulates exocrine pancreatic secretion in dogs and rats. Pancreas 2: 414-421, 1987. PMID: 2442749.
- Hou Y, Ernst SA, Heidenreich K, and Williams JA. Glucagon-like peptide-1 receptor is present in pancreatic acinar cells and regulates amylase secretion through cAMP. Am J Physiol Gastrointest Liver Physiol 310: G26-33, 2016. PMID: 26542397.
- Huang Y, Fischer JE, and Balasubramaniam A. Amylin mobilizes [Ca2+]i and stimulates the release of pancreatic digestive enzymes from rat acinar AR42J cells: evidence for an exclusive receptor system of amylin. Peptides 17: 497-502, 1996. PMID: 8735978.
- Iijima F, Iwatsuki K, and Chiba S. Effects of dopamine on exocrine secretion and cyclic nucleotide concentration in the dog pancreas. Eur J Pharmacol 92: 191-197, 1983. PMID: 6313394.
- Imoto I, Yamamoto M, Jia DM, and Otsuki M. Effect of chronic oral administration of the CCK receptor antagonist loxiglumide on exocrine and endocrine pancreas in normal rats. Int J Pancreatol 22: 177-185, 1997. PMID: 9444548.
- Inoue K, Hosotani R, Tatemoto K, Yajima H, and Tobe T. Effect of natural peptide YY on blood flow and exocrine secretion of pancreas in dogs. Dig Dis Sci 33: 828-832, 1988. PMID: 3378477.
- Iwatsuki K, Horiuchi A, Ren LM, and Chiba S. D-1 dopamine receptors mediate dopamine-induced pancreatic exocrine secretion in anesthetized dogs. Hypertens Res 18: S173-174, 1995. PMID: 8529053.
- Iwatsuki K, Horiuchi A, Ren LM, and Chiba S. Direct and indirect stimulation of pancreatic exocrine secretion by neurotensin in anaesthetized dogs. Clin Exp Pharmacol Physiol 18: 475-481, 1991. PMID: 1914248.
- Iwatsuki K, Ren LM, and Chiba S. Effects of peptide histidine isoleucine on pancreatic exocrine secretion in anaesthetized dogs. Clin Exp Pharmacol Physiol 20: 501-507, 1993. PMID: 8403531.
- Jaworek J, Bonior J, Konturek SJ, Bilski J, Szlachcic A, and Pawlik WW. Role of leptin in the control of postprandial pancreatic enzyme secretion. J Physiol Pharmacol 54: 591-602, 2003. PMID: 14726613.
- Jaworek J, Leja-Szpak A, Nawrot-Porabka K, Szklarczyk J, Kot M, Pierzchalski P, Goralska M, Ceranowicz P, Warzecha Z, Dembinski A, and Bonior J. Effects of Melatonin and Its Analogues on Pancreatic Inflammation, Enzyme Secretion, and Tumorigenesis. Int J Mol Sci 18, 2017. PMID: 28481310.
- Jaworek J, Nawrot-Porabka K, Leja-Szpak A, Bonior J, Szklarczyk J, Kot M, Konturek SJ, and Pawlik WW. Melatonin as modulator of pancreatic enzyme secretion and pancreatoprotector. J Physiol Pharmacol 58 Suppl 6: 65-80, 2007. PMID: 18212401.
- Jaworek J, Nawrot K, Konturek SJ, Leja-Szpak A, Thor P, and Pawlik WW. Melatonin and its precursor, L-tryptophan: influence on pancreatic amylase secretion in vivo and in vitro. J Pineal Res 36: 155-164, 2004. PMID: 15009505.
- Jaworek J, Szklarczyk J, Jaworek AK, Nawrot-Porabka K, Leja-Szpak A, Bonior J, and Kot M. Protective effect of melatonin on acute pancreatitis. Int J Inflam 2012: 173675, 2012. PMID: 22606640.
- Jensen SL, Holst JJ, Nielsen OV, and Rehfeld JF. Effect of sulfation of CCK-8 on its stimulation of the endocrine and exocrine secretion from the isolated perfused porcine pancreas. Digestion 22: 305-309, 1981. PMID: 6277714.
- Ji B, Bi Y, Simeone D, Mortensen RM, and Logsdon CD. Human pancreatic acinar cells lack functional responses to cholecystokinin and gastrin. Gastroenterology 121: 1380-1390, 2001. PMID: 11729117.
- Jin H, Cai L, Lee K, Chang TM, Li P, Wagner D, and Chey WY. A physiological role of peptide YY on exocrine pancreatic secretion in rats. Gastroenterology 105: 208-215, 1993. PMID: 8514036.
- Joehl RJ and DeJoseph MR. Pancreatic polypeptide inhibits amylase release by rat pancreatic acini. J Surg Res 40: 310-314, 1986. PMID: 2422440.
- Johnson CL, Weston JY, Chadi SA, Fazio EN, Huff MW, Kharitonenkov A, Köester A, and Pin CL. Fibroblast growth factor 21 reduces the severity of cerulein-induced pancreatitis in mice. Gastroenterology 137: 1795-1804, 2009. PMID: 19664632.
- Jowell PS, Robuck-Mangum G, Mergener K, Branch MS, Purich ED, and Fein SH. A double-blind, randomized, dose response study testing the pharmacological efficacy of synthetic porcine secretin. Aliment Pharmacol Ther 14: 1679-1684, 2000. PMID: 11121918.
- Kaelberer MM, Buchanan KL, Klein ME, Barth BB, Montoya MM, Shen X, and Bohorquez DV. A gut-brain neural circuit for nutrient sensory transduction. Science 361, 2018. PMID: 30237325.
- Kane MA, Kelley K, Ross SE, and Portanova LB. Isolation of a gastrin releasing peptide receptor from normal rat pancreas. Peptides 12: 207-213, 1991. PMID: 1648710.
- Kashimura J, Shimosegawa T, Kikuchi Y, Yoshida K, Koizumi M, Mochizuki T, Yanaihara N, and Toyota T. Effects of galanin on amylase secretion from dispersed rat pancreatic acini. Pancreas 9: 258-262, 1994. PMID: 7514793.
- Katoh K, Murai K, and Nonoyama T. Effects of substance P on fluid and amylase secretion in exocrine pancreas of rat and mouse. Res Vet Sci 36: 147-152, 1984. PMID: 6201966.
- Katschinski M, Dahmen G, Reinshagen M, Beglinger C, Koop H, Nustede R, and Adler G. Cephalic stimulation of gastrointestinal secretory and motor responses in humans. Gastroenterology 103: 383-391, 1992. PMID: 1634057.
- Kawabata A, Kuroda R, Nishida M, Nagata N, Sakaguchi Y, Kawao N, Nishikawa H, Arizono N, and Kawai K. Protease-activated receptor-2 (PAR-2) in the pancreas and parotid gland: Immunolocalization and involvement of nitric oxide in the evoked amylase secretion. Life Sci 71: 2435-2446, 2002. PMID: 12231404.
- Kayasseh L, Haecki WH, Gyr K, Stalder GA, Rittmann WW, Halter F, and Girard J. The endogenous release of pancreatic polypeptide by acid and meal in dogs. Effect of somatostatin. Scand J Gastroenterol 13: 385-391, 1978. PMID: 675146.
- Kemeny LV, Hegyi P, Rakonczay Z, Jr., Borka K, Korompay A, Gray MA, Argent BE, and Venglovecz V. Substance P inhibits pancreatic ductal bicarbonate secretion via neurokinin receptors 2 and 3 in the guinea pig exocrine pancreas. Pancreas 40: 793-795, 2011. PMID: 21673544.
- Kikuchi Y, Koizumi M, Shimosegawa T, Kashimura J, Suzuki S, and Toyota T. Islet amyloid polypeptide has no effect on amylase release from rat pancreatic acini stimulated by CCK-8, secretin, carbachol and VIP. Tohoku J Exp Med 165: 41-48, 1991. PMID: 1724710.
- Kim C, Kim K, Lee H, Song C, Ryu H, and Hyun J. Potentiation of cholecystokinin and secretin-induced pancreatic exocrine secretion by endogenous insulin in humans. Pancreas 18: 410-414, 1999. PMID: 10231848.
- Kim CK, Lee KY, Wang T, Sun G, Chang TM, and Chey WY. Role of endogenous cholecystokinin on vagally stimulated pancreatic secretion in dogs. Am J Physiol 257: G944-949, 1989. PMID: 2481981.
- Klein E, Salinas A, Shemesh E, and Dreiling DA. Effects of autonomic denervation on canine exocrine pancreatic secretion and blood flow. Int J Pancreatol 3: 165-170, 1988. PMID: 3361158.
- Knuhtsen S, Holst JJ, Jensen SL, Knigge U, and Nielsen OV. Gastrin-releasing peptide: effect on exocrine secretion and release from isolated perfused porcine pancreas. Am J Physiol 248: G281-286, 1985. PMID: 3976887.
- Koevary SB, McEvoy RC, and Azmitia EC. Specific uptake of tritiated serotonin in the adult rat pancreas: evidence for the presence of serotonergic fibers. Am J Anat 159: 361-368, 1980. PMID: 6971051.
- Koh YH and Bhatia M. Substance P. Pancreapedia: Exocrine Pancreas Knowledge Base, 2011. DOI: 10.3998/panc.2011.23.
- Kohler E, Beglinger C, Eysselein V, Grotzinger U, and Gyr K. Gastrin is not a physiological regulator of pancreatic exocrine secretion in the dog. Am J Physiol 252: G40-44, 1987. PMID: 3812686.
- Kohler H, Nustede R, Barthel, M, Müller C, Schafmayer A Total denervation of the pancreas does not alter the pancreatic polypeptide-induced inhibition of pancreatic exocrine secretion in dogs. Res Exp Med (Berl) 191: 359-369, 1991. PMID: 8446771.
- Konturek JW, Gabryelewicz A, Kulesza E, Konturek SJ, and Domschke W. Cholecystokinin (CCK) in the amino acid uptake and enzyme protein secretion by the pancreas in humans. Int J Pancreatol 17: 55-61, 1995. PMID: 8568335.
- Konturek SJ, Bielanski W, and Solomon TE. Effects of an antral mucosectomy, L-364,718 and atropine on cephalic phase of gastric and pancreatic secretion in dogs. Gastroenterology 98: 47-55, 1990. PMID: 2403431.
- Konturek SJ, Dubiel J, and Gabrys B. Effect of acid infusion into various levels of the intestine on gastric and pancreatic secretion in the cat. Gut 10: 749-753, 1969. PMID: 5386632.
- Konturek SJ, Meyers CA, Kwiecien N, Obtulowicz W, Tasler J, Oleksy J, Kopp B, Coy DH, and Schally AV. Effect of human pancreatic polypeptide and its C-terminal hexapeptide on pancreatic secretion in man and in the dog. Scand J Gastroenterol 17: 395-399, 1982. PMID: 7134866.
- Konturek SJ, Radecki T, Mikos E, and Thor J. The effect of exogenous and endogenous secretin and cholecystokinin on pancreatic secretion in cats. Scand J Gastroenterol 6: 423-428, 1971. PMID: 5093529.
- Konturek SJ, Tasler J, Bilski J, Cieszkowski M, Cai RZ, and Schally AV. Antagonism of receptors for gastrin, cholecystokinin and GRP/bombesin in postprandial stimulation of exocrine pancreas in dogs. J Physiol Pharmacol 44: 43-53, 1993. PMID: 8390873.
- Konturek SJ, Tasler J, Cieszkowski M, Jaworek J, and Konturek J. Intravenous amino acids and fat stimulate pancreatic secretion. Am J Physiol 236: E678-684, 1979. PMID: 443422.
- Konturek SJ, Tasler J, Cieszkowski M, Szewczyk K, and Hladij M. Effect of cholecystokinin receptor antagonist on pancreatic responses to exogenous gastrin and cholecystokinin and to meal stimuli. Gastroenterology 94: 1014-1023, 1988. PMID: 3345871.
- Konturek SJ, Yanaihara N, Pawlik W, Jaworek J, and Szewczyk K. Comparison of helodermin, VIP and PHI in pancreatic secretion and blood flow in dogs. Regul Pept 24: 155-166, 1989. PMID: 2922493.
- Koop I, Schindler M, Bosshammer A, Scheibner J, Stange E, and Koop H. Physiological control of cholecystokinin release and pancreatic enzyme secretion by intraduodenal bile acids. Gut 39: 661-667, 1996. PMID: 9026479.
- Kopin AS, Mathes WF, McBride EW, Nguyen M, Al-Haider W, Schmitz F, Bonner-Weir S, Kanarek R, and Beinborn M. The cholecystokinin-A receptor mediates inhibition of food intake yet is not essential for the maintenance of body weight. J Clin Invest 103: 383-391, 1999. PMID: 9927499.
- Kreiss C, Schwizer W, Erlacher U, Borovicka J, Lochner-Kuery C, Muller R, Jansen JB, and Fried M. Role of antrum in regulation of pancreaticobiliary secretion in humans. Am J Physiol 270: G844-851, 1996. PMID: 8967497.
- Kuvshinoff BW, Brodish RJ, James L, McFadden DW, and Fink AS. Somatostatin inhibits secretin-induced canine pancreatic response via a cholinergic mechanism. Gastroenterology 105: 539-547, 1993. PMID: 8101500.
- Lai KC, Cheng CH, and Leung PS. The ghrelin system in acinar cells: localization, expression, and regulation in the exocrine pancreas. Pancreas 35: e1-8, 2007. PMID: 17895831.
- Larsson LI and Rehfeld JF. Peptidergic and adrenergic innervation of pancreatic ganglia. Scand J Gastroenterol 14: 433-437, 1979. PMID: 384501.
- Lee E, Gerlach U, Uhm DY, and Kim J. Inhibitory effect of somatostatin on secretin-induced augmentation of the slowly activating K+ current (IKs) in the rat pancreatic acinar cell. Pflugers Arch 443: 405-410, 2002. PMID: 11810210.
- Lee KY, Krusch D, Zhou L, Song Y, Chang TM, and Chey WY. Effect of endogenous insulin on pancreatic exocrine secretion in perfused dog pancreas. Pancreas 11: 190-195, 1995. PMID: 7479678.
- Lee KY, Lee YL, Kim CD, Chang TM, and Chey WY. Mechanism of action of insulin on pancreatic exocrine secretion in perfused rat pancreas. Am J Physiol 267: G207-212, 1994. PMID: 7915495.
- Lee W, Miyazaki K, and Funakoshi A. Effects of somatostatin and pancreatic polypeptide on exocrine and endocrine pancreas in the rats. Gastroenterol Jpn 23: 49-55, 1988. PMID: 2895031.
- Lee YL, Kwon HY, Park HS, Lee TH, and Park HJ. The role of insulin in the interaction of secretin and cholecystokinin in exocrine secretion of the isolated perfused rat pancreas. Pancreas 12: 58-63, 1996. PMID: 8927620.
- Leja-Szpak A, Jaworek J, Nawrot-Porabka K, Palonek M, Mitis-Musiol M, Dembinski A, Konturek SJ, and Pawlik WW. Modulation of pancreatic enzyme secretion by melatonin and its precursor; L-tryptophan. Role of CCK and afferent nerves. J Physiol Pharmacol 55 Suppl 2: 33-46, 2004. PMID: 15608359.
- Lenz HJ, Messmer B, and Zimmerman FG. Noradrenergic inhibition of canine gallbladder contraction and murine pancreatic secretion during stress by corticotropin-releasing factor. J Clin Invest 89: 437-443, 1992. PMID: 1737835.
- Lhoste E, Aprahamian M, Pousse A, Hoeltzel A, and Stock-Damge C. Combined effect of chronic bombesin and secretin or cholecystokinin on the rat pancreas. Peptides 6 Suppl 3: 83-87, 1985. PMID: 2421268.
- Lhoste EF, Levenez F, Chabanet C, Fiszlewicz M, and Corring T. Effect of bombesin at low doses on the secretion of the exocrine pancreas and on plasma gastrin concentration in the conscious pig. Regul Pept 74: 41-45, 1998. PMID: 9657358.
- Liang T, Dolai S, Xie L, Winter E, Orabi AI, Karimian N, Cosen-Binker LI, Huang YC, Thorn P, Cattral MS, and Gaisano HY. Ex vivo human pancreatic slice preparations offer a valuable model for studying pancreatic exocrine biology. J Biol Chem 292: 5957-5969, 2017. PMID: 28242761.
- Liddle RA. Regulation of cholecystokinin secretion by intraluminal releasing factors. Am J Physiol Gastrointest Liver Physiol 269: G319-G327, 1995. PMID: 7573441.
- Liddle RA. Regulation of Pancreatic Secretion. In: Physiology of the Gastrointestinal Tract. 5th Ed. Vol 2. Edited by Johnson LR, et al. Academic Press, 2012. p. 1425-1460. DOI: 10.1016/ B978-0-12-382026-6.00052-X
- Liddle RA, Goldfine ID, Rosen MS, Taplitz RA, and Williams JA. Cholecystokinin bioactivity in human plasma. Molecular forms, responses to feeding, and relationship to gallbladder contraction. J Clin Invest 75: 1144-1152, 1985. PMID: 2580857.
- Liddle RA, Green GM, Conrad CK, and Williams JA. Proteins but not amino acids, carbohydrates, or fats stimulate cholecystokinin secretion in the rat. Am J Physiol 251: G243-248, 1986. PMID: 3740265.
- Liddle RA, Misukonis MA, Pacy L, and Balber AE. Cholecystokinin cells purified by fluorescence-activated cell sorting respond to monitor peptide with an increase in intracellular calcium. Proc Natl Acad Sci USA 89: 5147-5151, 1992. PMID: 1594624.
- Liddle RA, Rushakoff RJ, Morita ET, Beccaria L, Carter JD, and Goldfine ID. Physiological role for cholecystokinin in reducing postprandial hyperglycemia in humans. J Clin Invest 81: 1675-1681, 1988. PMID: 3290250.
- Lin TM, Evans DC, Shaar CJ, and Root MA. Action of somatostatin on stomach, pancreas, gastric mucosal blood flow, and hormones. Am J Physiol 244: G40-45, 1983. PMID: 6129805.
- Liou AP, Lu X, Sei Y, Zhao X, Pechhold S, Carrero RJ, Raybould HE, and Wank S. The G-protein-coupled receptor GPR40 directly mediates long-chain fatty acid-induced secretion of cholecystokinin. Gastroenterology 140: 903-912, 2011. PMID: 20955703.
- Liou AP, Sei Y, Zhao X, Feng J, Lu X, Thomas C, Pechhold S, Raybould HE, and Wank SA. The extracellular calcium-sensing receptor is required for cholecystokinin secretion in response to L-phenylalanine in acutely isolated intestinal I cells. Am J Physiol Gastrointest Liver Physiol 300: G538-546, 2011. PMID: 21252045.
- Liu HP, Tay SS, and Leong SK. Nitrergic neurons in the pancreas of newborn guinea pig: their distribution and colocalization with various neuropeptides and dopamine-beta-hydroxylase. J Auton Nerv Syst 61: 248-256, 1996. PMID: 8988482.
- Lopez J and Cuesta N. Adrenomedullin as a pancreatic hormone. Microsc Res Tech 57: 61-75, 2002. PMID: 11921357.
- Louie DS, Williams JA, and Owyang C. Action of pancreatic polypeptide on rat pancreatic secretion: in vivo and in vitro. Am J Physiol 249: G489-495, 1985. PMID: 2413769.
- Love JA and Szebeni K. Morphology and histochemistry of the rabbit pancreatic innervation. Pancreas 18: 53-64, 1999. PMID: 9888661.
- Maack T. Role of atrial natriuretic factor in volume control. Kidney Int 49: 1732-1737, 1996. PMID: 8743487.
- MacGregor I, Parent J, and Meyer JH. Gastric emptying of liquid meals and pancreatic and biliary secretion after subtotal gastrectomy or truncal vagotomy and pyloroplasty in man. Gastroenterology 72: 195-205, 1977. PMID: 830568.
- Malagelada JR, DiMagno EP, Summerskill WH, and Go VL. Regulation of pancreatic and gallbladder functions by intraluminal fatty acids and bile acids in man. J Clin Invest 58: 493-499, 1976. PMID: 956380.
- Malagelada JR, Go VL, DiMagno EP, and Summerskill WH. Interactions between intraluminal bile acids and digestive products on pancreatic and gallbladder function. J Clin Invest 52: 2160-2165, 1973. PMID: 4727454.
- Mannon P and Taylor IL. The pancreatic polypeptide family. In: Gut peptides: Biochemistry and Physiology. Edited by Walsh JH and Dockray GJ. New York: Raven Press. p: 341-370. 1994.
- Marongiu L, Perra MT, Pinna AD, Sirigu F, and Sirigu P. Peptidergic (VIP) nerves in normal human pancreas and in pancreatitis: an immunohistochemical study. Histol Histopathol 8: 127-132, 1993. PMID: 8443423.
- Matsushita K, Okabayashi Y, Koide M, Hasegawa H, Otsuki M, and Kasuga M. Potentiating effect of insulin on exocrine secretory function in isolated rat pancreatic acini. Gastroenterology 106: 200-206, 1994. PMID: 7506218.
- Matyjek R, Herzig KH, Kato S, and Zabielski R. Exogenous leptin inhibits the secretion of pancreatic juice via a duodenal CCK1-vagal-dependent mechanism in anaesthetized rats. Regul Pept 114: 15-20, 2003. PMID: 12763635.
- Mayer EA, Thompson JB, Jehn D, Reedy T, Elashoff J, and Meyer JH. Gastric emptying and sieving of solid food and pancreatic and biliary secretion after solid meals in patients with truncal vagotomy and antrectomy. Gastroenterology 83: 184-192, 1982. PMID: 6919504.
- McLaughlin J. Long-chain fatty acid sensing in the gastrointestinal tract. Biochem Soc Trans 35: 1199-1202, 2007. PMID: 17956311.
- McLaughlin J, Grazia Luca M, Jones MN, D'Amato M, Dockray GJ, and Thompson DG. Fatty acid chain length determines cholecystokinin secretion and effect on human gastric motility. Gastroenterology 116: 46-53, 1999. PMID: 9869601.
- McLaughlin JT, Lomax RB, Hall L, Dockray GJ, Thompson DG, and Warhurst G. Fatty acids stimulate cholecystokinin secretion via an acyl chain length-specific, Ca2+-dependent mechanism in the enteroendocrine cell line STC-1. J Physiol 513: 11-18, 1998. PMID: 9782155.
- Messmer B, Zimmerman FG, and Lenz HJ. Regulation of exocrine pancreatic secretion by cerebral TRH and CGRP: role of VIP, muscarinic, and adrenergic pathways. Am J Physiol 264: G237-242, 1993. PMID: 8447406.
- Meuth-Metzinger VL, Philouze-Rome V, Metzinger L, Gespach C, and Guilloteau P. Differential activation of adenylate cyclase by secretin and VIP receptors in the calf pancreas. Pancreas 31: 174-181, 2005. PMID: 16025005.
- Meyer JH and Grossman MI. Comparison of D- and L-phenylalanine as pancreatic stimulants. Am J Physiol 222: 1058-1063, 1972. PMID: 5027085.
- Meyer JH and Jones RS. Canine pancreatic responses to intestinally perfused fat and products of fat digestion. Am J Physiol 226: 1178-1187, 1974. PMID: 4824870.
- Meyer JH and Kelly GA. Canine pancreatic responses to intestinally perfused proteins and protein digests. Am J Physiol 231: 682-691, 1976. PMID: 970450.
- Meyer JH, Kelly GA, and Jones RS. Canine pancreatic response to intestinally perfused oligopeptides. Am J Physiol 231: 678-681, 1976. PMID: 970449.
- Meyer JH, Kelly GA, Spingola LJ, and Jones RS. Canine gut receptors mediating pancreatic responses to luminal L-amino acids. Am J Physiol 231: 669-677, 1976. PMID: 970448.
- Meyer JH, Spingola J, and Grossman MI. Endogenous cholecystokinin potentiates exogenous secretin on pancreas of dog. Am J Physiol 221: 742-747, 1971. PMID: 5570331.
- Meyer JH, Way LW, and Grossman MI. Pancreatic bicarbonate response to various acids in duodenum of the dog. Am J Physiol 219: 964-970, 1970. PMID: 5459498.
- Migita Y, Nakano I, Goto M, Ito T, and Nawata H. Effect of pancreastatin on cerulein-stimulated pancreatic blood flow and exocrine secretion in anaesthetized rats. J Gastroenterol Hepatol 14: 583-587, 1999. PMID: 10385069.
- Miller TA, Watson LC, Rayford PL, and Thompson JC. The effect of glucagon on pancreatic secretion and plasma secretin in dogs. World J Surg 1: 93-97, 1977. PMID: 17233.
- Miyasaka K, Funakoshi A, Nakamura R, Kitani K, Shimizu F, and Tatemoto K. Effects of porcine pancreastatin on postprandial pancreatic exocrine secretion and endocrine functions in the conscious rat. Digestion 43: 204-211, 1989. PMID: 2612743.
- Miyasaka K, Funakoshi A, Shikado F, and Kitani K. Stimulatory and inhibitory effects of bile salts on rat pancreatic secretion. Gastroenterology 102: 598-604, 1992. PMID: 1343077.
- Miyasaka K, Funakoshi A, Yasunami Y, Nakamura R, Kitani K, Tamamura H, Funakoshi S, and Fujii N. Rat pancreastatin inhibits both pancreatic exocrine and endocrine secretions in rats. Regul Pept 28: 189-198, 1990. PMID: 1693005.
- Miyasaka K and Green GM. Effect of partial exclusion of pancreatic juice on rat basal pancreatic secretion. Gastroenterology 86: 114-119, 1984. PMID: 6689654.
- Miyasaka K and Kitani K. A difference in stimulatory effects on pancreatic exocrine secretion between ursodeoxycholate and trypsin inhibitor in the rat. Dig Dis Sci 31: 978-986, 1986. PMID: 2426065.
- Miyasaka K and Kitani K. Effects of bile salts on pancreatic secretion in rabbits: ursodeoxycholate infused into the duodenum stimulates pancreas. Pancreas 1: 264-269, 1986. PMID: 3575308.
- Miyasaka K, Sazaki N, Funakoshi A, Matsumoto M, and Kitani K. Two mechanisms of inhibition by bile on luminal feedback regulation of rat pancreas. Gastroenterology 104: 1780-1785, 1993. PMID: 8500737.
- Miyasaka K, Shinozaki H, Jimi A, and Funakoshi A. Amylase secretion from dispersed human pancreatic acini: neither cholecystokinin a nor cholecystokinin B receptors mediate amylase secretion in vitro. Pancreas 25: 161-165, 2002. PMID: 12142739.
- Miyasaka K, Shinozaki H, Suzuki S, Sato Y, Kanai S, Masuda M, Jimi A, Nagata A, Matsui T, Noda T, Kono A, and Funakoshi A. Disruption of cholecystokinin (CCK)-B receptor gene did not modify bile or pancreatic secretion or pancreatic growth: a study in CCK-B receptor gene knockout mice. Pancreas 19: 114-118, 1999. PMID: 10438156.
- Moghimzadeh E, Ekman R, Hakanson R, Yanaihara N, and Sundler F. Neuronal gastrin-releasing peptide in the mammalian gut and pancreas. Neuroscience 10: 553-563, 1983. PMID: 6355896.
- Moore EW, Verine HJ, and Grossman MI. Pancreatic bicarbonate response to a meal. Acta Hepatogastroenterol (Stuttg) 26: 30-36, 1979. PMID: 34310.
- Morisset J, Levenez F, Corring T, Benrezzak O, Pelletier G, and Calvo E. Pig pancreatic acinar cells possess predominantly the CCK-B receptor subtype. Am J Physiol 271: E397-402, 1996. PMID: 8770035.
- Moriyasu M, Lee YL, Lee KY, Chang TM, and Chey WY. Effect of digested protein on pancreatic exocrine secretion and gut hormone release in the dog. Pancreas 9: 129-133, 1994. PMID: 8108365.
- Mulholland MW, Lally K, and Taborsky GJ, Jr. Inhibition of rat pancreatic exocrine secretion by neuropeptide Y: studies in vivo and in vitro. Pancreas 6: 433-440, 1991. PMID: 1715087.
- Müller TD, Finan B, Bloom SR, D'Alessio D, Drucker DJ, Flatt PR, Fritsche A, Gribble F, Grill HJ, Habener JF, Holst JJ, Langhans W, Meier JJ, Nauck MA, Perez-Tilve D, Pocai A, Reimann F, Sandoval DA, Schwartz TW, Seeley RJ, Stemmer K, Tang-Christensen M, Woods SC, DiMarchi RD, and Tschöp MH. Glucagon-like peptide 1 (GLP-1). Mol Metab 30: 72-130, 2019. PMID: 31767182.
- Mulvihill SJ, Bunnett NW, Goto Y, and Debas HT. Somatostatin inhibits pancreatic exocrine secretion via a neural mechanism. Metabolism 39: 143-148, 1990. PMID: 1698248.
- Murphy JA, Criddle DN, Sherwood M, Chvanov M, Mukherjee R, McLaughlin E, Booth D, Gerasimenko JV, Raraty MG, Ghaneh P, Neoptolemos JP, Gerasimenko OV, Tepikin AV, Green GM, Reeve JR, Jr., Petersen OH, and Sutton R. Direct activation of cytosolic Ca2+ signaling and enzyme secretion by cholecystokinin in human pancreatic acinar cells. Gastroenterology 135: 632-641, 2008. PMID: 18555802.
- Mussa BM, Sartor DM, and Verberne AJM. Dorsal vagal preganglionic neurons: Differential responses to CCK1 and 5-HT3 receptor stimulation. Auton Neurosci 156: 36-43, 2010. PMID: 20346737.
- Mussa BM and Verberne AJ. Activation of the dorsal vagal nucleus increases pancreatic exocrine secretion in the rat. Neurosci Lett 433: 71-76, 2008. PMID: 18241988.
- Nakajima S, Hira T, and Hara H. Calcium-sensing receptor mediates dietary peptide-induced CCK secretion in enteroendocrine STC-1 cells. Mol Nutr Food Res 56: 753-760, 2012. PMID: 22648622.
- Navas JM, Calvo JJ, Lopez MA, and De Dios I. Exocrine pancreatic response to intraduodenal fatty acids and fats in rabbits. Comp Biochem Physiol Comp Physiol 105: 141-145, 1993. PMID: 8099868.
- Nawrot-Porabka K, Jaworek J, Leja-Szpak A, Palonek M, Szklarczyk J, Konturek SJ, and Pawlik WW. Leptin is able to stimulate pancreatic enzyme secretion via activation of duodeno-pancreatic reflex and CCK release. J Physiol Pharmacol 55 Suppl 2: 47-57, 2004. PMID: 15608360.
- Nawrot-Porabka K, Jaworek J, Leja-Szpak A, Szklarczyk J, Konturek SJ, and Reiter RJ. Luminal melatonin stimulates pancreatic enzyme secretion via activation of serotonin-dependent nerves. Pharmacol Rep 65: 494-504, 2013. PMID: 23744434.
- Nawrot-Porabka K, Jaworek J, Leja-Szpak A, Szklarczyk J, Kot M, Mitis-Musiol M, Konturek SJ, and Pawlik WW. Involvement of vagal nerves in the pancreatostimulatory effects of luminal melatonin, or its precursor L-tryptophan. Study in the rats. J Physiol Pharmacol 58 Suppl 6: 81-95, 2007. PMID: 18212402.
- Niebergall-Roth E and Singer MV. Central and peripheral neural control of pancreatic exocrine secretion. J Physiol Pharmacol 52: 523-538, 2001. PMID: 11787756.
- Niederau C, Sonnenberg A, and Erckenbrecht J. Effects of intravenous infusion of amino acids, fat, or glucose on unstimulated pancreatic secretion in healthy humans. Dig Dis Sci 30: 445-455, 1985. PMID: 2580678.
- Niijima A. Effect of umami taste stimulations on vagal efferent activity in the rat. Brain Res Bull 27: 393-396, 1991. PMID: 1959036.
- Nishimura T, Nakatake Y, Konishi M, and Itoh N. Identification of a novel FGF, FGF-21, preferentially expressed in the liver. Biochim Biophys Acta 1492: 203-206, 2000. PMID: 10858549.
- Nustede R, Schmidt WE, Kohler H, Folsch UR, and Schafmayer A. The influence of bile acids on the regulation of exocrine pancreatic secretion and on the plasma concentrations of neurotensin and CCK in dogs. Int J Pancreatol 13: 23-30, 1993. PMID: 8454915.
- Oguchi H, Iwatsuki K, Horiuchi A, Furuta S, and Chiba S. Effects of human atrial natriuretic polypeptide on pancreatic exocrine secretion in the dog. Biochem Biophys Res Commun 146: 757-763, 1987. PMID: 2956951.
- Ohnishi H, Mine T, and Kojima I. Inhibition by somatostatin of amylase secretion induced by calcium and cyclic AMP in rat pancreatic acini. Biochem J 304: 531-536, 1994. PMID: 7528010.
- Okabayashi Y, Otsuki M, Ohki A, Sakamoto C, and Baba S. Effects of C-terminal fragments of cholecystokinin on exocrine and endocrine secretion from isolated perfused rat pancreas. Endocrinology 113: 2210-2215, 1983. PMID: 6196184.
- Olsen O, Schaffalitzky de Muckadell OB, and Cantor P. Fat and pancreatic secretion. Scand J Gastroenterol 24: 74-80, 1989. PMID: 2928726.
- Osnes M, Hanssen LE, Flaten O, and Myren J. Exocrine pancreatic secretion and immunoreactive secretin (IRS) release after intraduodenal instillation of bile in man. Gut 19: 180-184, 1978. PMID: 631638.
- Osnes M, Hanssen LE, Lehnert P, Flaten O, Larsen S, Londong W, and Otte M. Exocrine pancreatic secretion and immunoreactive secretin release after repeated intraduodenal infusions of bile in man. Scand J Gastroenterol 15: 1033-1039, 1980. PMID: 6165067.
- Otsuki M, Okabayashi Y, Ohki A, Oka T, Fujii M, Nakamura T, Sugiura N, Yanaihara N, and Baba S. Action of cholecystokinin analogues on exocrine and endocrine rat pancreas. Am J Physiol 250: G405-411, 1986. PMID: 2421584.
- Pandol SJ. The Exocrine Pancreas. San Rafael (CA): Morgan & Claypool Life Sciences, 2010. PMID: 21634067.
- Pandol SJ, Schoeffield MS, Sachs G, and Muallem S. Role of free cytosolic calcium in secretagogue-stimulated amylase release from dispersed acini from guinea pig pancreas. J Biol Chem 260: 10081-10086, 1985. PMID: 2410419.
- Pandol SJ, Sutliff VE, Jones SW, Charlton CG, O'Donohue TL, Gardner JD, and Jensen RT. Action of natural glucagon on pancreatic acini: due to contamination by previously undescribed secretagogues. Am J Physiol 245: G703-710, 1983. PMID: 6195929.
- Pandol SJ, Thomas MW, Schoeffield MS, Sachs G, and Muallem S. Role of calcium in cholecystokinin-stimulated phosphoinositide breakdown in exocrine pancreas. Am J Physiol 248: G551-560, 1985. PMID: 2581457.
- Pappas TN, Debas HT, Goto Y, and Taylor IL. Peptide YY inhibits meal-stimulated pancreatic and gastric secretion. Am J Physiol 248: G118-123, 1985. PMID: 3838121.
- Pappas TN, Debas HT, and Taylor IL. Peptide YY: metabolism and effect on pancreatic secretion in dogs. Gastroenterology 89: 1387-1392, 1985. PMID: 3840443.
- Pariente JA, Madrid JA, and Salido GM. Role of histamine receptors in rabbit pancreatic exocrine secretion stimulated by cholecystokinin and secretin. Exp Physiol 75: 657-667, 1990. PMID: 1978780.
- Park HJ, Lee YL, and Kwon HY. Effects of pancreatic polypeptide on insulin action in exocrine secretion of isolated rat pancreas. J Physiol 463: 421-429, 1993. PMID: 7504106.
- Park HS, Kwon HY, Lee YL, Chey WY, and Park HJ. Role of GRPergic neurons in secretin-evoked exocrine secretion in isolated rat pancreas. Am J Physiol Gastrointest Liver Physiol 278: G557-562, 2000. PMID: 10762609.
- Park HS, Lee YL, Kwon HY, Chey WY, and Park HJ. Significant cholinergic role in secretin-stimulated exocrine secretion in isolated rat pancreas. Am J Physiol 274: G413-418, 1998. PMID: 9486197.
- Park YD, Cui ZY, and Park HJ. Effects of gamma-aminobutyric acid on action of gastrin-releasing peptidergic neurons in exocrine secretion of isolated, perfused rat pancreas. Pancreas 25: 308-313, 2002. PMID: 12370544.
- Patel R, Singh J, Yago MD, Vilchez JR, Martinez-Victoria E, and Manas M. Effect of insulin on exocrine pancreatic secretion in healthy and diabetic anaesthetised rats. Mol Cell Biochem 261: 105-110, 2004. PMID: 15362492.
- Pearson GT, Singh J, and Petersen OH. Adrenergic nervous control of cAMP-mediated amylase secretion in the rat pancreas. Am J Physiol 246: G563-573, 1984. PMID: 6202157.
- Phillips PA, Yang L, Shulkes A, Vonlaufen A, Poljak A, Bustamante S, Warren A, Xu Z, Guilhaus M, Pirola R, Apte MV, and Wilson JS. Pancreatic stellate cells produce acetylcholine and may play a role in pancreatic exocrine secretion. Proc Natl Acad Sci USA 107: 17397-17402, 2010. PMID: 20852067.
- Pisegna JR, de Weerth A, Huppi K, and Wank SA. Molecular cloning of the human brain and gastric cholecystokinin receptor: structure, functional expression and chromosomal localization. Biochem Biophys Res Commun 189: 296-303, 1992. PMID: 1280419.
- Powers MA, Schiffman SS, Lawson DC, Pappas TN, and Taylor IL. The effect of taste on gastric and pancreatic responses in dogs. Physiol Behav 47: 1295-1297, 1990. PMID: 2395936.
- Reeve JR, Jr., Green GM, Chew P, Eysselein VE, and Keire DA. CCK-58 is the only detectable endocrine form of cholecystokinin in rat. Am J Physiol Gastrointest Liver Physiol 285: G255-265, 2003. PMID: 12686511.
- Riepl RL, Fiedler F, Kowalski C, Teufel J, and Lehnert P. Exocrine pancreatic secretion and plasma levels of cholecystokinin, pancreatic polypeptide, and somatostatin after single and combined intraduodenal application of different bile salts in man. Ital J Gastroenterol 28: 421-429, 1996. PMID: 9032583.
- Riepl RL, Fiedler F, Teufel J, and Lehnert P. Effect of intraduodenal bile and taurodeoxycholate on exocrine pancreatic secretion and on plasma levels of vasoactive intestinal polypeptide and somatostatin in man. Pancreas 9: 109-116, 1994. PMID: 7509061.
- Riepl RL and Lehnert P. The role of bile in the regulation of exocrine pancreatic secretion. Scand J Gastroenterol 27: 625-631, 1992. PMID: 1439544.
- Riepl RL, Reichardt B, Rauscher J, Tzavella K, Teufel J, and Lehnert P. Mediators of exocrine pancreatic secretion induced by intraduodenal application of bile and taurodeoxycholate in man. Eur J Med Res 2: 23-29, 1997. PMID: 9049590.
- Rodríguez MR, Diez F, Ventimiglia MS, Morales V, Copsel S, Vatta MS, Davio CA, and Bianciotti LG. Atrial Natriuretic Factor Stimulates Efflux of cAMP in Rat Exocrine Pancreas via Multidrug Resistance–Associated Proteins. Gastroenterology 140: 1292-1302, 2011. PMID: 21237168.
291. Rothman SS. Molecular regulation of digestion: short term and bond specific. Am J Physiol 226: 77-83, 1974. PMID: 4809890.
292. Rune SJ. pH in the human duodenum. Its physiological and pathophysiological significance. Digestion 8: 261-268, 1973. PMID: 4721705.
293. Sabbatini ME, Bi Y, Ji B, Ernst SA, and Williams JA. CCK activates RhoA and Rac1 differentially through Gα13 and Gαq in mouse pancreatic acini. Am J Physiol Cell Physiol 298: C592-C601, 2010. PMID: 19940064.
294. Sabbatini ME, Rodriguez M, di Carlo MB, Davio CA, Vatta MS, and Bianciotti LG. C-type natriuretic peptide enhances amylase release through NPR-C receptors in the exocrine pancreas. Am J Physiol Gastrointest Liver Physiol 293: G987-994, 2007. PMID: 17702953.
295. Sabbatini ME, Rodriguez MR, Dabas P, Vatta MS, and Bianciotti LG. C-type natriuretic peptide stimulates pancreatic exocrine secretion in the rat: role of vagal afferent and efferent pathways. Eur J Pharmacol 577: 192-202, 2007. PMID: 17900562.
296. Sabbatini ME, Vatta MS, Davio CA, and Bianciotti LG. Atrial natriuretic factor negatively modulates secretin intracellular signaling in the exocrine pancreas. Am J Physiol Gastrointest Liver Physiol 292: G349-357, 2007. PMID: 16973919.
297. Sabbatini ME, Villagra A, Davio CA, Vatta MS, Fernandez BE, and Bianciotti LG. Atrial natriuretic factor stimulates exocrine pancreatic secretion in the rat through NPR-C receptors. Am J Physiol Gastrointest Liver Physiol 285: G929-937, 2003. PMID: 12829435.
298. Sakamoto C, Otsuki M, Ohki A, Yuu H, Maeda M, Yamasaki T, and Baba S. Glucose-dependent insulinotropic action of cholecystokinin and caerulein in the isolated perfused rat pancreas. Endocrinology 110: 398-402, 1982. PMID: 6173205.
299. Sanchez-Vicente C, Rodriguez-Nodal F, Minguela A, Garcia LJ, San Roman JI, Calvo JJ, and Lopez MA. Cholinergic pathways are involved in secretin and VIP release and the exocrine pancreatic response after intraduodenally perfused acetic and lactic acids in the rat. Pancreas 10: 93-99, 1995. PMID: 7899466.
300. Sans MD, Sabbatini ME, Ernst SA, D'Alecy LG, Nishijima I, and Williams JA. Secretin is not necessary for exocrine pancreatic development and growth in mice. Am J Physiol Gastrointest Liver Physiol 301: G791-G798, 2011. PMID: 21852360.
301. Sarles H, Dani R, Prezelin G, Souville C, and Figarella C. Cephalic phase of pancreatic secretion in man. Gut 9: 214-221, 1968. PMID: 5655031.
302. Satoh K, Ouchi M, Morita A, and Kashimata M. MARCKS phosphorylation and amylase release in GLP-1-stimulated acini isolated from rat pancreas. J Physiol Sci 69: 143-149, 2019. PMID: 29845509.
303. Schaffalitzky de Muckadell OB, Fahrenkrug J, Matzen P, Rune SJ, and Worning H. Physiological significance of secretin in the pancreatic bicarbonate secretion. II. Pancreatic bicarbonate response to a physiological increase in plasma secretin concentration. Scand J Gastroenterol 14: 85-90, 1979. PMID: 424692.
304. Schaffalitzky de Muckadell OB, Fahrenkrug J, Watt-Boolsen S, and Worning H. Pancreatic response and plasma secretin concentration during infusion of low dose secretin in man. Scand J Gastroenterol 13: 305-311, 1978. PMID: 755275.
305. Schafmayer A, Nustede R, Pompino A, and Kohler H. Vagal influence on cholecystokinin and neurotensin release in conscious dogs. Scand J Gastroenterol 23: 315-320, 1988. PMID: 3387897.
306. Schmid R, Schulte-Frohlinde E, Schusdziarra V, Neubauer J, Stegmann M, Maier V, and Classen M. Contribution of postprandial amino acid levels to stimulation of insulin, glucagon, and pancreatic polypeptide in humans. Pancreas 7: 698-704, 1992. PMID: 1448457.
307. Schwartz TW. Pancreatic polypeptide: a hormone under vagal control. Gastroenterology 85: 1411-1425, 1983. PMID: 6138294.
308. Schwartz TW, Stenquist B, and Olbe L. Cephalic phase of pancreatic-polypeptide secretion studied by sham feeding in man. Scand J Gastroenterol 14: 313-320, 1979. PMID: 441680.
309. Seifert H, Sawchenko P, Chesnut J, Rivier J, Vale W, and Pandol SJ. Receptor for calcitonin gene-related peptide: binding to exocrine pancreas mediates biological actions. Am J Physiol 249: G147-151, 1985. PMID: 2409816.
310. Sheikh SP, Holst JJ, Skak-Nielsen T, Knigge U, Warberg J, Theodorsson-Norheim E, Hokfelt T, Lundberg JM, and Schwartz TW. Release of NPY in pig pancreas: dual parasympathetic and sympathetic regulation. Am J Physiol 255: G46-54, 1988. PMID: 3389414.
311. Sheikh SP, Roach E, Fuhlendorff J, and Williams JA. Localization of Y1 receptors for NPY and PYY on vascular smooth muscle cells in rat pancreas. Am J Physiol 260: G250-257, 1991. PMID: 1847590.
312. Shimosegawa T, Asakura T, Kashimura J, Yoshida K, Meguro T, Koizumi M, Mochizuki T, Yanaihara N, and Toyota T. Neurons containing gastrin releasing peptide-like immunoreactivity in the human pancreas. Pancreas 8: 403-412, 1993. PMID: 8361958.
313. Shimosegawa T, Moriizumi S, Koizumi M, Kashimura J, Yanaihara N, and Toyota T. Immunohistochemical demonstration of galaninlike immunoreactive nerves in the human pancreas. Gastroenterology 102: 263-271, 1992. PMID: 1370157.
314. Shiratori K, Watanabe S, and Takeuchi T. Inhibitory effect of intraduodenal administration of somatostatin analogue SDZ CO 611 on rat pancreatic exocrine secretion. Pancreas 8: 471-475, 1993. PMID: 8361968.
315. Shiratori K, Watanabe S, and Takeuchi T. Somatostatin analog, SMS 201-995, inhibits pancreatic exocrine secretion and release of secretin and cholecystokinin in rats. Pancreas 6: 23-30, 1991. PMID: 1704631.
316. Singer M. Neurohormonal Control of Pancreatic Enzyme Secretion in Animals. In: The Pancreas: Biology, Pathobiology, and Disease. 2nd ed. Edited by Go VLW, et al. New York: Raven Press. p. 425-448. 1993.
317. Singer MV. Pancreatic secretory response to intestinal stimulants: a review. Scand J Gastroenterol Suppl 139: 1-13, 1987. PMID: 3324297.
318. Singer MV and Niebergall-Roth E. Secretion from acinar cells of the exocrine pancreas: role of enteropancreatic reflexes and cholecystokinin. Cell Biol Int 33: 1-9, 2009. PMID: 18948215.
319. Singer MV, Solomon TE, and Grossman MI. Effect of atropine on secretion from intact and transplanted pancreas in dog. Am J Physiol 238: G18-22, 1980. PMID: 6986796.
320. Singer MV, Solomon TE, Rammert H, Caspary F, Niebel W, Goebell H, and Grossman MI. Effect of atropine on pancreatic response to HCl and secretin. Am J Physiol 240: G376-380, 1981. PMID: 7235024.
321. Singer MV, Tiscornia OM, Mendes de Oliveiro JP, Demol P, Levesque D, and Sarles H. Effect of glucagon on canine exocrine pancreatic secretion stimulated by a test meal. Can J Physiol Pharmacol 56: 1-6, 1978. PMID: 638847.
322. Singh J, Pariente JA, and Salido GM. The physiological role of histamine in the exocrine pancreas. Inflamm Res 46: 159-165, 1997. PMID: 9197985.
323. Singh J and Pearson GT. Effects of nerve stimulation on enzyme secretion from the in vitro rat pancreas and 3H-release after preincubation with catecholamines. Naunyn Schmiedebergs Arch Pharmacol 327: 228-233, 1984. PMID: 6208490.
324. Singh M. Effect of glucagon on digestive enzyme synthesis, transport and secretion in mouse pancreatic acinar cells. J Physiol 306: 307-322, 1980. PMID: 6162027.
325. Solomon TE. Regulation of pancreatic secretion. Clin Gastroenterol 13: 657-678, 1984. PMID: 6386236.
326. Solomon TE, Solomon N, Shanbour LL, and Jacobson ED. Direct and indirect effects of nicotine on rabbit pancreatic secretion. Gastroenterology 67: 276-283, 1974. PMID: 4847707.
327. Sommer H and Kasper H. The action of synthetic secretin, cholecystokinin-octapeptide and combinations of these hormones on the secretion of the isolated perfused rat pancreas. Hepatogastroenterology 28: 311-315, 1981. PMID: 6178671.
328. Spannagel AW, Green GM, Guan D, Liddle RA, Faull K, and Reeve JR, Jr. Purification and characterization of a luminal cholecystokinin-releasing factor from rat intestinal secretion. Proc Natl Acad Sci USA 93: 4415-4420, 1996. PMID: 8633081.
329. Stabile BE, Borzatta M, Stubbs RS, and Debas HT. Intravenous mixed amino acids and fats do not stimulate exocrine pancreatic secretion. Am J Physiol 246: G274-280, 1984. PMID: 6322595.
330. Stock-Damge C, Lhoste E, Aprahamian M, and Loza E. Influence of repeated administration of bombesin on rat pancreatic secretion. Pancreas 2: 658-663, 1987. PMID: 3438303.
331. Sumi S, Inoue K, and Tobe T. Experimental studies on the interrelationship between organs mediated by peptide YY: effect on splanchnic circulation and exocrine pancreas in dogs. Nihon Geka Hokan 59: 224-233, 1990. PMID: 2130785.
332. Szalmay G, Varga G, Kajiyama F, Yang XS, Lang TF, Case RM, and Steward MC. Bicarbonate and fluid secretion evoked by cholecystokinin, bombesin and acetylcholine in isolated guinea-pig pancreatic ducts. J Physiol 535: 795-807, 2001. PMID: 11559776.
333. Takahashi M, Naito H, Sasaki I, Funayama Y, Shibata C, and Matsuno S. Long-term bile diversion enhances basal and duodenal oleate-stimulated pancreatic exocrine secretion in dogs. Tohoku J Exp Med 203: 87-95, 2004. PMID: 15212143.
334. Takiguchi S, Suzuki S, Sato Y, Kanai S, Miyasaka K, Jimi A, Shinozaki H, Takata Y, Funakoshi A, Kono A, Minowa O, Kobayashi T, and Noda T. Role of CCK-A receptor for pancreatic function in mice: a study in CCK-A receptor knockout mice. Pancreas 24: 276-283, 2002. PMID: 11893936.
335. Tang C, Biemond I, and Lamers CB. Cholecystokinin receptors in human pancreas and gallbladder muscle: A comparative study. Gastroenterology 111: 1621-1626, 1996. PMID: 8942742.
336. Tang C, Biemond I, and Lamers CB. Visualization and characterization of CCK receptors in exocrine pancreas of rat with storage phosphor autoradiography. Pancreas 13: 311-315, 1996. PMID: 8884854.
337. Tatemoto K. Isolation and characterization of peptide YY (PYY), a candidate gut hormone that inhibits pancreatic exocrine secretion. Proc Natl Acad Sci USA 79: 2514-2518, 1982. PMID: 6953409.
338. Taylor IL, Byrne WJ, Christie DL, Ament ME, and Walsh JH. Effect of individual l-amino acids on gastric acid secretion and serum gastrin and pancreatic polypeptide release in humans. Gastroenterology 83: 273-278, 1982. PMID: 6806140.
339. Teyssen S, Niebergall E, Chari ST, and Singer MV. Comparison of two dose-response techniques to study the pancreatic secretory response to intraduodenal tryptophan in the absence and presence of the M1-receptor antagonist telenzepine. Pancreas 10: 368-373, 1995. PMID: 7792293.
340. Thimister PW, Hopman WP, Tangerman A, Rosenbusch G, Willems HL, and Jansen JB. Effect of intraduodenal bile salt on pancreaticobiliary responses to bombesin and to cholecystokinin in humans. Hepatology 28: 1454-1460, 1998. PMID: 9828206.
341. Trimble ER, Bruzzone R, Biden TJ, and Farese RV. Secretin induces rapid increases in inositol trisphosphate, cytosolic Ca2+ and diacylglycerol as well as cyclic AMP in rat pancreatic acini. Biochem J 239: 257-261, 1986. PMID: 3028367.
342. Tsuchida T, Ohnishi H, Tanaka Y, Mine T, and Fujita T. Inhibition of stimulated amylase secretion by adrenomedullin in rat pancreatic acini. Endocrinology 140: 865-870, 1999. PMID: 9927317.
343. Tu HJ, Zhao CF, Chen ZW, Lin W, and Jiang YC. Fibroblast Growth Factor (FGF) Signaling Protects Against Acute Pancreatitis-Induced Damage by Modulating Inflammatory Responses. Med Sci Monit 26: e920684, 2020. PMID: 32283546.
344. Turkelson CM, Solomon TE, Bussjaeger L, Turkelson J, Ronk M, Shively JE, Ho FJ, and Reeve JR, Jr. Chemical characterization of rat cholecystokinin-58. Peptides 9: 1255-1260, 1988. PMID: 3247248.
345. Ulrich CD, 2nd, Wood P, Hadac EM, Kopras E, Whitcomb DC, and Miller LJ. Cellular distribution of secretin receptor expression in rat pancreas. Am J Physiol 275: G1437-1444, 1998. PMID: 9843782.
346. Vagne M and Grossman MI. Gastric and pancreatic secretion in response to gastric distention in dogs. Gastroenterology 57: 300-310, 1969. PMID: 4897231.
347. Van Dijk A, Richards JG, Trzeciak A, Gillessen D, and Mohler H. Cholecystokinin receptors: biochemical demonstration and autoradiographical localization in rat brain and pancreas using [3H] cholecystokinin8 as radioligand. J Neurosci 4: 1021-1033, 1984. PMID: 6325601.
348. Varga G, Papp M, and Vizi ES. Cholinergic and adrenergic control of enzyme secretion in isolated rat pancreas. Dig Dis Sci 35: 501-507, 1990. PMID: 1690631.
349. Ventimiglia MS, Najenson AC, Rodríguez MR, Davio CA, Vatta MS, and Bianciotti LG. Natriuretic Peptides and Their Receptors. In: The Pancreapedia: Exocrine Pancreas Knowledge Base, 2011. DOI: 10.3998/panc.2011.36.
350. von Schonfeld J, Muller MK, Runzi M, Geling M, Neisius I, Kleimann J, and Goebell H. Pancreastatin--a mediator in the islet-acinar axis? Metabolism 42: 552-555, 1993. PMID: 8492708.
351. Wang Y, Chandra R, Samsa LA, Gooch B, Fee BE, Cook JM, Vigna SR, Grant AO, and Liddle RA. Amino acids stimulate cholecystokinin release through the Ca2+-sensing receptor. Am J Physiol Gastrointest Liver Physiol 300: G528-537, 2011. PMID: 21183662.
352. Wheeler S, Eardley JE, McNulty KF, Sutcliffe CP, and Morrison JD. An investigation into the relative merits of pituitary adenylate cyclase-activating polypeptide (PACAP-27) and vasoactive intestinal polypeptide as vagal neuro-transmitters in exocrine pancreas of rats. Exp Physiol 82: 729-747, 1997. PMID: 9257115.
353. White TT, Lundh G, and Magee DF. Evidence for the existence of a gastropancreatic reflex. Am J Physiol 198: 725-728, 1960. PMID: 13844402.
354. White TT, Mc AR, and Magee DF. Gastropancreatic reflex after various gastric operations. Surg Forum 13: 286-288, 1962. PMID: 14000361.
355. Williams JA. Bombesin. In: The Pancreapedia: Exocrine Pancreas Knowledge Base, 2015. DOI: 10.3998/panc.2015.10.
356. Williams JA. GLP-1. In: The Pancreapedia: Exocrine Pancreas Knowledge Base, 2014. DOI: 10.3998/panc.2014.7.
357. Williams JA. Pancreatic Polypeptide. In: The Pancreapedia: Exocrine Pancreas Knowledge Base, 2014. DOI: 10.3998/panc.2014.4.
358. Williams JA. Secretin. In: The Pancreapedia: Exocrine Pancreas Knowledge Base, 2013. DOI: 10.3998/panc.2013.5.
359. Williams JA, Chen X, and Sabbatini ME. Small G proteins as key regulators of pancreatic digestive enzyme secretion. Am J Physiol Endocrinol Metab 296: E405-414, 2009. PMID: 19088252.
360. Wolfe BM, Keltner RM, and Kaminski DL. The effect of an intraduodenal elemental diet on pancreatic secretion. Surg Gynecol Obstet 140: 241-245, 1975. PMID: 1124475.
361. Yago MD, Manas M, Ember Z, and Singh J. Nitric oxide and the pancreas: morphological base and role in the control of the exocrine pancreatic secretion. Mol Cell Biochem 219: 107-120, 2001. PMID: 11354241.
362. Yago MD, Tapia JA, Salido GM, Adeghate E, Juma LM, Martinez-Victoria E, Manas M, and Singh J. Effect of sodium nitroprusside and 8-bromo cyclic GMP on nerve-mediated and acetylcholine-evoked secretory responses in the rat pancreas. Br J Pharmacol 136: 49-56, 2002. PMID: 11976267.
363. Yamaguchi N and Fukushima Y. PACAP enhances stimulation-induced norepinephrine release in canine pancreas in vivo. Can J Physiol Pharmacol 76: 788-797, 1998. PMID: 10030460.
364. Yi E, Smith TG, Baker RC, and Love JA. Catecholamines and 5-hydroxytryptamine in tissues of the rabbit exocrine pancreas. Pancreas 29: 218-224, 2004. PMID: 15367888.
365. You CH, Rominger JM, and Chey WY. Effects of atropine on the action and release of secretin in humans. Am J Physiol 242: G608-611, 1982. PMID: 7091334.
366. You CH, Rominger JM, and Chey WY. Potentiation effect of cholecystokinin-octapeptide on pancreatic bicarbonate secretion stimulated by a physiologic dose of secretin in humans. Gastroenterology 85: 40-45, 1983. PMID: 6303892.
367. Young AA, Jodka C, Pittner R, Parkes D, and Gedulin BR. Dose-response for inhibition by amylin of cholecystokinin-stimulated secretion of amylase and lipase in rats. Regul Pept 130: 19-26, 2005. PMID: 15982756.
368. Zhang W, Chen M, Chen X, Segura BJ, and Mulholland MW. Inhibition of pancreatic protein secretion by ghrelin in the rat. J Physiol 537: 231-236, 2001. PMID: 11711576.