
Entry Version:
Citation:
Pancreapedia: Exocrine Pancreas Knowledge Base, DOI: 10.3998/panc.2020.06
I. Introduction
The human exocrine pancreas secretes 1-2 liters of pancreatic juice each day. When stimulated, the pancreas secretes alkaline pancreatic juice containing copious amounts of bicarbonate (HCO3-) (27, 86). HCO3- plays essential roles in the digestive system. HCO3- determines the pH of bodily fluids as the major buffer system that guards against toxic pH fluctuations (133). HCO3− in pancreatic juice neutralizes gastric acid, and provides an optimal pH environment for digestive enzymes to function in the duodenum (86). In addition, HCO3− acts as a moderate chaotropic ion that facilitates the solubilization of macromolecules, such as digestive enzymes and mucins (47). The importance of pancreatic HCO3- secretion is highlighted in the abnormal HCO3- secretion in several forms of pancreatitis (118, 168) and in cystic fibrosis (CF), which causes poor mucin hydration and solubilization leading to obstruction of ductal structures of the pancreas, intestine, vas deferens and the lung (129, 130).
The exocrine pancreas is composed of three major cell types, acinar, duct and stellate cells. Acinar cells secrete a small volume of isotonic, plasma-like, NaCl-rich fluid and digestive enzymes. Duct cells modify the ionic composition of the fluid and secrete the bulk of the fluid and HCO3−of the pancreatic juice. Stellate cells may aid the pancreas recovery from injury (82). As the main HCO3- secretor, the duct has key roles in the development of acute and chronic pancreatitis. At pH 7.4 and 5% CO2, the HCO3- concentration in plasma is approximately 25 mM. In human, dog, cat, and guinea pig, HCO3- concentration in postprandial pancreatic juice is higher than 140 mM (27, 86). This remarkable transport performance has attracted much attention from pancreatologists and physiologists. Current understanding of the molecular mechanism of pancreatic HCO3- secretion was improved by the recent identification of ion transporters and channels, including the cystic fibrosis transmembrane conductance regulator (CFTR) (69), the electrogenic Na+-HCO3- co-transporter NBCe1-B (also known an pNBC1) (1), and the solute-linked carrier 26 (SLC26) transporters (29, 116), together with regulatory proteins, such as with-no-lysine kinase 1 (WNK1) (119), STE20/SPS1-related proline/alanine-rich kinase (SPAK) (35) and the inositol-1,4,5-triphosphate (IP3) receptor binding protein released with IP3 (IRBIT) (163) and their role in pancreatic HCO3- secretion.
II. Control of Pancreatic HCO3- Secretion
Pancreatic HCO3- secretion increases in response to ingestion of a meal and is regulated by multiple neurohumoral inputs. Fluid and enzyme secretion by acinar cells are controlled predominantly by an increase in cytoplasmic free Ca2+ concentration ([Ca2+]i) (103, 123, 124). Fluid and HCO3- secretion by duct cells are regulated by the second messengers cAMP (86, 101) that synergizes with Ca2+ to generate the physiological response (4, 97, 122). Pancreatic ductal cells express receptors for a battery of hormones and neurotransmitters. The two major hormones controlling pancreatic fluid and HCO3- secretion are the Gs-coupled, cAMP generating hormone secretin and the Gq-coupled, Ca2+ mobilizing hormone cholecystokinin (CCK), which are released from gastrointestinal endocrine cells in the upper duodenum. Cholinergic vagal output via an enteropancreatic vagovagal reflex also has an important role in controlling ductal fluid and HCO3- secretion. In addition to these classic stimuli, several other humoral agents are released by the pancreas for fine tuning its secretion, including insulin, somatostatin, purines, and prostaglandins (90). Additional information on hormonal control of pancreatic secretion can be found in a previous review (86) and the “Regulation of Pancreatic Secretion” section in Pancreapedia (21).
A. Humoral Control
Secretin
The low pH (below 4.5) of gastric chyme stimulates the release of secretin from duodenal S cells into the blood (15, 22). Secretin stimulates ductal fluid and HCO3- secretion and synergizes with Ca2+ mobilizing agonists to potentiate enzyme secretion by acinar cells. Plasma secretin levels rise after a meal (22, 127) and correlate with HCO3- output (135). Secretin-stimulated fluid and HCO3- secretion is modulated directly or indirectly by both peptide hormones, such as CCK and somatostatin, and by vagal stimulation (43, 77, 167).
CCK
CCK is a major stimulator of acinar cell enzyme and fluid secretion which is mediated by the Ca2+-dependent exocytosis of zymogen granules and activation of apical (luminal) Cl- channels, respectively. The synaptotagmins are the Ca2+ sensor that convey the Ca2+ signal for pancreatic exocytosis (104) and Ca2+ activates the Ca2+-activated Anoctamin 1 (TMEM16A) to initiates acinar cells fluid secretion (117). CCK also acts on pancreatic duct secretion; however, the effects of CCK on pancreatic duct differ among species. In humans, the effect of CCK alone on ductal fluid secretion is weak; however, CCK greatly potentiates the effects of secretin (167).
Purines
Pancreatic duct cells express multiple purinergic type 2 receptor (P2Rs) types, including ionotropic P2X and metabotropic P2Y receptors at the apical and basolateral membranes (96). P2Rs are stimulated by purinergic ligands released from nerve terminals at the basolateral space, zymogen granules of acinar cells into the luminal space, or efflux by ductal ATP transporters to both the basolateral and luminal compartments (79). Stimulation of P2Rs increases [Ca2+]i in duct cells (112, 114). Several studies have examined effects of P2Rs on ion transporters in ductal cell lines, but there are almost no studies on ductal fluid and HCO3- secretion. Ishiguro et al. demonstrated that luminal ATP stimulated, while basolateral ATP inhibited fluid and HCO3- secretion in guinea-pig pancreatic duct (57). More recent studies examined the effect of various stimuli and ion channels of ATP release from ductal cell lines (79) that will be important to verify in native ducts.
B. Neuronal Control
Pancreatic secretion is regulated by the enteric nervous system, which is composed of a gut-brain axis and an intrapancreatic system. The major neurotransmitter acting on pancreatic duct cells is acetylcholine released by vagal parasympathetic fibers. Duct cells express both M1 and M3 muscarinic receptors, which act through changes in [Ca2+]i. The M3 receptors maybe more prominent based on their higher expression level relative to the M1 receptors (36, 71). In humans, cholinergic stimulation enhances ductal secretion stimulated by secretin, likely by synergistic mechanism that is mediated by IRBIT (4, 122). Vasoactive intestinal peptide (VIP) and ATP are also localized in parasympathetic nerve terminals (78, 113). Vagal stimulation causes VIP release that is associated with fluid and HCO3- secretion (52, 64, 78).
III. Key Transporters Involved in Pancreatic HCO3- Secretion
Pancreatic HCO3- secretion is mediated by a coordinated function of transporters expressed in the apical and basolateral membranes of duct cells. Pancreatic HCO3- secretion can be divided into 2 steps. The first step is uptake of HCO3- into duct cells from the blood through the basolateral membrane. The second step is efflux of HCO3- across the apical membrane of duct cells. Regulatory mechanisms in the cytosol that include ions like Cl- and several kinases and phosphatases, act on the transporters to coordinate and integrate the secretory process. Recent advances in molecular, cellular, and physiological techniques have enhanced our understanding of the molecular identity, localization, function, and regulatory mechanisms of ductal ion transporters (4, 87, 97). The major ion transporters expressed in the apical and basolateral membranes of the pancreatic duct cells are summarized in Table 1 and Figure 1.
Table 1: Transporters of pancreatic duct
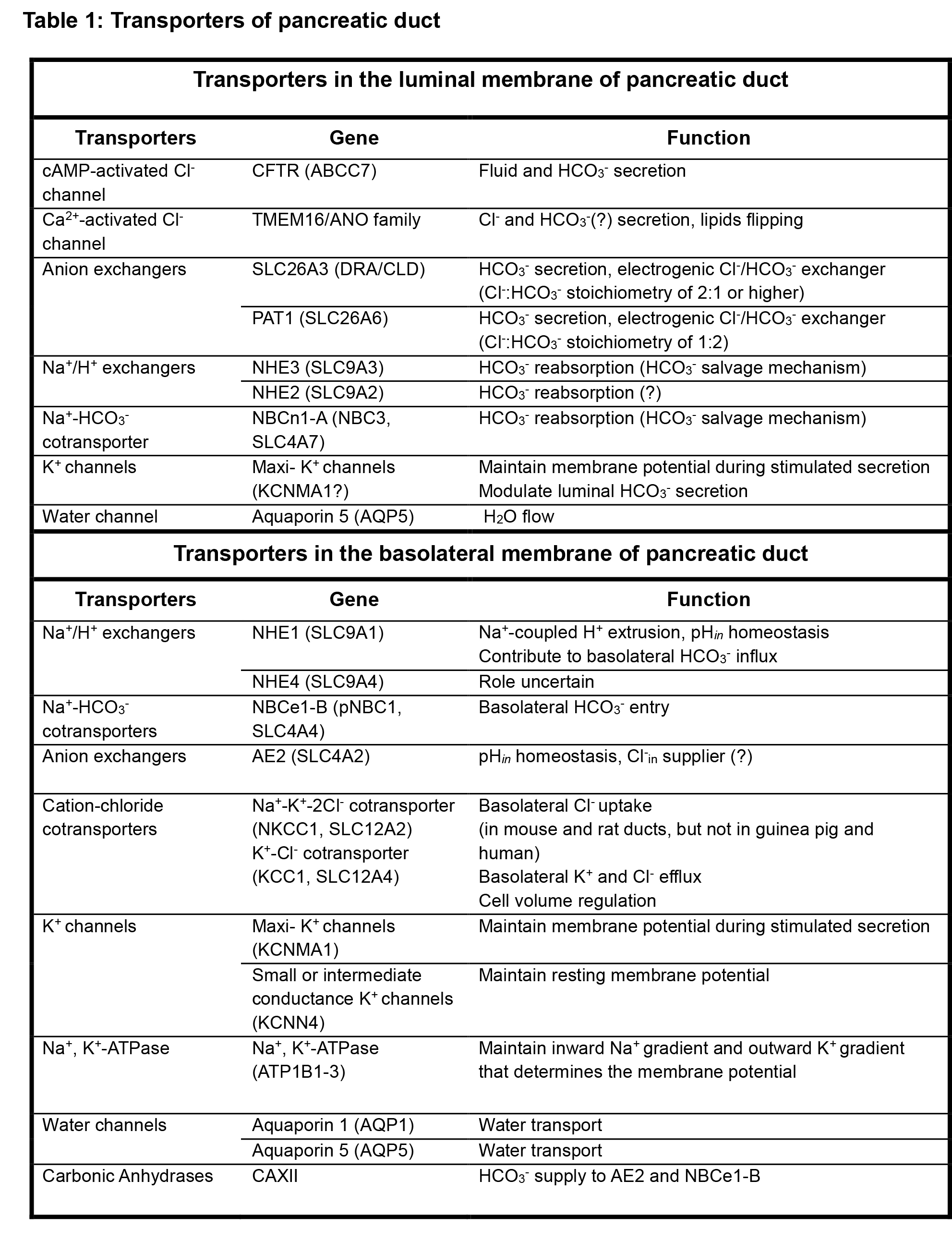
Figure 1. A schematic diagram depicting the transporters and channels in the apical (luminal) and basolateral membranes of pancreatic duct cells. The main driving force for HCO3- secretion is achieved by the Na+ gradient generated by the Na+/K+ ATPase pump and K+ channels at the basolateral membrane, which generate the intracellular negative membrane potential. HCO3- is loaded mainly through the electrogenic (1Na+-2HCO3-) NBCe1-B, and partly by NHE1 located in the basolateral membrane. Basolateral AE2 may act to supply Cl-in to maintain the secretion. Apical HCO3- secretion is performed by the interacting and functionally interrelated CFTR and Slc26a6. Transcellular HCO3- movement generates a lumen-negative electrical potential that results in paracellular Na+ secretion through the paracellular pathway. Water follows Na+ and HCO3- osmotically via paracellular and transcellular (aquaporins) pathways. In the resting state, luminal NHE3 and NBCn1-A function as salvage luminal HCO3-. Modified from (87).
A. Na+/K+ ATPase, and K+ Channels
The main driving force for fluid secretion is achieved by the Na+/K+ ATPase pump and K+ channels which generate the transmembrane Na+ and K+ gradients and the negative intracellular membrane potential (87, 118). The Na+/K+ ATPase pump is expressed in the basolateral membrane of the ducts (99, 134, 145, 150), and generates the Na+ and K+ gradients by extruding 3 Na+ ions in exchange for uptake of 2 extracellular K+ ions using the energy of ATP hydrolysis. K+ channels in both the basolateral and apical membranes use the K+ gradient generated by the pump to generate a negative membrane potential. The Na+ gradient is used to drive several Na+-coupled solutes, including HCO3- absorption by the basolateral Na+-HCO3- cotransporter NBCe1-B and basolateral and luminal Na+/H+ exchangers (NHEs). The negative membrane potential aids in controlling HCO3- uptake by NBCe1-B and in HCO3- efflux through luminal electrogenic transporters. MaxiK channels (KCNMA1) have been identified on the basolateral membrane of rat pancreatic duct cells, and are likely candidates for maintaining the membrane potential during agonist-stimulated HCO3- secretion (38). A Ba2+-sensitive channel of 82 pS conductance (KCNN4) appears to be a basolateral K+ channel, which is responsible for the resting K+ permeability (115). Apical membrane K+ channels were identified in acinar cells (5) and in pancreatic duct cells, with the later having a role in ductal HCO3- secretion (155).
B. Na+-HCO3- Co-transporters (NBCs)
The main ductal basolateral membrane HCO3- accumulation transporter is NBCe1-B (87). NBCe1-B was cloned from pancreas and was named pNBC1 (1). It was later re-named NBCe1-B as part of classification of the NBC family (14). NBCe1-B is an electrogenic transporter with a 1 Na+: 2 HCO3- stoichiometry in pancreatic duct cells (42). NBCe1-B can be regulated by cAMP-dependent protein kinase A (PKA) phosphorylation at Ser1026 and Thr49 (41). In principle, Na+/H+ exchangers in the basolateral membrane (e.g. NHE1) can also mediate HCO3- influx in duct cells. However, the electrogenic NBCe1-B utilizes the Na+ gradient more efficiently than the electroneutral NHE1 (1 Na+: 1 HCO3-). Indeed, NBCe1-B contributes up to ~75% of the HCO3- influx during secretin-induced ductal fluid and HCO3- secretion in guinea pig (60, 62). The activity of NBCe1-B is controlled by multiple inputs, including IRBIT and the WNK/Ste20-related proline/alanine-rich kinase (SPAK) pathway (143, 164) and most notably intracellular Cl- (138). A more recent analysis revealed an intricate regulation of NBCe1-B by the WNK and CaMKII (Ca2+ and calmodulin activated kinase II) kinases and the SPAK and calcineurin phosphatases the dephosphorylate the serine residues phosphorylated by the respective kinases (153). The kinases/phosphatases pairs determine regulation of NBCe1-B by Cl-in (153), which emerges as a new general form of signaling ion (97). In addition to NBCe1-B, the duct expresses electroneutral NBCn1-A (NBC3) on the apical (luminal) membrane (120, 128). This transporter may mediate HCO3- salvage in the resting state to maintain acidified pancreatic juice (37, 100).
C. CFTR
The discovery of acidic pancreatic juice in patients with cystic fibrosis (CF) was a milestone in understanding the mechanism of pancreatic HCO3- secretion (65, 81). The CF transmembrane conductance regulator (CFTR) was discovered as the protein mutated in patients with CF (69, 131, 132). Although CFTR is a member of the ATP-binding cassette (ABC) transporter superfamily that usually act as membrane pumps that transport their substrates against the electrochemical gradient (24), CFTR functions as an anion (Cl- and HCO3-) channel, through which ions diffuse down the electrochemical gradient. CFTR is located at the apical membrane of pancreatic ducts (20, 148, 170) (and all secretory epithelia), and is activated by the cAMP/PKA pathway. At [Cl-]i higher than 10 mM, CFTR functions as a Cl- channel that has limited permeability to HCO3- (92, 126, 137). However, when [Cl-]i drops to below 10 mM, CFTR anionic selectivity changes to increase HCO3- permeability and mediate luminal HCO3- exit (67, 119). Indeed, as has been shown in patients with CF (20, 56, 148), CFTR is critically involved in epithelial HCO3- secretion. This leads to revision of the original model of ductal HCO3- secretion, in which Cl-/HCO3- exchangers mediate apical HCO3- efflux and CFTR facilitates the apical Cl-/HCO3- exchangers by recycling the Cl- (12). This continues to be the case at high Cl-in. However, at low [Cl-]i, HCO3- efflux via CFTR driven by the membrane potential has essential role in HCO3- efflux and HCO3--driven fluid secretion in the pancreatic duct (61, 147). The dynamic change in CFTR Cl-/HCO3- permeability is mediated by the protein kinase WNK1 (70, 119). WNK1 (125) and other members of the WNK kinases are regulated by Cl-in, with high Cl-in in the low (WNK4) and high (WNK1) range inhibiting the WNKs. Reduction in Cl-in activates the WNKs that act directly or through SPAK on CFTR and other HCO3-, Na+, Cl- and K+ transporters (97). To regulate their activity and selectivity. If is of interest that the WNKs show differential sensitivity to Cl-in and effect on the transporters (151, 163). Thus, a modest reduction in Cl-in is sufficient to activate WNK1 and increase HCO3- transport by CFTR (119). Further reduction in Cl-in will activate WNK4 that inhibits CFTR activity (163), perhaps to prevent excess HCO3- secretion, that is energetically very expensive involving transport by multiple electrogenic transporters. The significance of CFTR-dependent HCO3- secretion in CFTR-expressing epithelia, including the pancreas, has been established in a study correlating CFTR-dependent HCO3- transport and severity of the CF disease (23). The importance of the shift in the WNK1-mediated shift in CFTR HCO3- selectivity has been clearly demonstrated in a study that examined pancreatitis-associated CFTR mutations with altered WNK1-mediated increase in HCO3- permeability and found clear correlation between reduced HCO3- permeability and chronic pancreatitis in humans (83).
CFTR has a more global role in ductal fluid and HCO3- secretion. In addition to functioning as a Cl- and HCO3- channel, CFTR functions as a scaffold forming macromolecular complexes with other transporters and regulatory proteins at the apical membrane (87). CFTR has a PSD95/Discs-large/ZO-1 (PDZ) ligand at the C-terminus and binds to PDZ domains of adapter proteins, such as Na+/H+ exchanger regulatory factors (NHERFs). It also has SH3 and multiple ankyrin repeat domains 2 (Shank2) (84, 144), through which CFTR interacts and regulates the activity of slc26a6, slc26a3 (75), NHE3 (3) and NBCn1-A (120). Other interactions of CFTR are with soluble NSF attachment protein receptor (SNARE) proteins, A-kinase anchor proteins (AKAPs), kinases and phosphatases (44) that may serve to regulate CFTR activity and the activity of the transporters interacting with CFTR. The interaction with the SLC26 transporters is of particular significance since the two transporters are mutually activated when interacting (75, 76). The mutual regulation is mediated by interaction of the CFTR R domain with the SLC26 transporters STAS domain (76).
D. Cl-/HCO3- Exchangers
Cl-/HCO3- exchangers mediate the bulk of HCO3- exit across the apical membranes of the pancreatic duct cells until the last portion of HCO3- exit that is mediated by CFTR once it gains HCO3- permeability. In humans, members of the solute-linked carrier 4 (SLC4) and the SLC26 families function as Cl-/HCO3- exchangers. Among the SLC4 transporters, duct cells express AE2 (SLC4A2) at the basolateral membrane that regulates pHi and protects against alkaline load (118). However, our studies revealed an essential role for AE2 in ductal fluid and HCO3- secretion (53). Intuitively, basolateral HCO3- efflux mechanism should inhibit rather than stimulate ductal HCO3- secretion. It is not clear why AE2 is essential for ductal fluid secretion. Maintaining stable pHin that neutralize acid load by the Na+/H+ exchangers and high pH next to the plasma membrane is one potential critical function of AE2. Another possibility is that AE2 may provide the duct with Cl- that is needed to keep the luminal slc26a6 functioning in a face of limited Cl- provided by acinar secretion (53).
Among the SLC26 family transporters, SLC26A3, and SLC26A6 are located on the apical membrane of the pancreatic duct cells and mediate Cl-/HCO3- exchange. Interestingly, SLC26A3 has a 2Cl-/1HCO3- stoichiometry (76, 139), while SLC26A6 functions as a 2HCO3-/1Cl- exchanger (72, 139). A persistent osmotic gradient is needed to support the copious fluid secretion by the pancreatic duct. This is satisfied by the coupled action of NBCe1-B and SLC26A6 that results in a continuous net HCO3- (osmolyte) transcellular transport and thus transcellular H2O flow (140, 149, 159). In addition, as indicated above, SLC26 transporters interact with CFTR through the sulfate transporter and anti-sigma factor antagonist (STAS) domain, and regulate pancreatic secretion by activating CFTR (76). This form of regulation is critical for pancreatic and other exocrine glands HCO3- secretion, including the pancreas, salivary glands, the kidney and the lung (91).
E. Other Transporters, Channels, and Carbonic Anhydrases
Na+/H+ exchangers (NHEs)
TheSLC9A NHE family contains electroneutral 1Na+/1H+ exchangers. The ubiquitous NHE1 (SLC9A1) is essential for intracellular pH homeostasis and supplies Na+ to the Na+/K+ ATPase pump on the basolateral membrane of the pancreatic duct (171). Diffusion of CO2 from the blood into the duct and CO2 generated by metabolism is hydrated by the action of carbonic anhydrases to generate HCO3- and H+. Consequently, H+ efflux by NHE1 may contribute to basolateral HCO3- uptake. However, NHE1 does not have a major role in basolateral HCO3- influx as revealed by minimal inhibition of fluid and HCO3- secretion by inhibition of NHE1 in pancreatic duct of most species (154, 162). The NHE3 isoform is expressed in the apical membrane of pancreatic duct and is thought to mediate HCO3- salvage at the resting state (85). At the resting state, the pancreatic juice is acidic, indicating an active H+ secretion (37, 100) that may be mediated by the combined action of NHE3 and NBCn1-A. Similar to NBCn1-A (120), NHE3 interacts with CFTR via PDZ domain containing proteins (3), and is regulated by IRBIT (48, 49). However, the physiological significance of these transporters await evaluation in mouse models with targeted pancreatic deletion of ductal NHEs and NBCn1-A.
Ca2+-activated Cl- channels (CaCCs)
Several members of the anoctamin (TMEM16/ANO) family function as CaCC (18, 136, 166). TMEM16A/ANO1, TMEM16B/ANO2, TMEM16F/ANO6, TMEM16H/ANO8, and TMEM16K/ANO10 are expressed in pancreas (87). However, ANO1 is expressed in acinar but not duct cells (136), ANO6 functions as a flipase and as a Cl- channel (150) and ANO8 is a tether at the ER/PM junctions that controls assembly of Ca2+ signaling complexes (63). The function of ANO2 and ANO10 in the pancreas is not clear at this time. Nevertheless, ample evidence shows that the pancreatic duct (and ducts of other secretory glands) has CaCC activity in the apical membrane (39, 40, 157, 169). The molecular identity of this channel is not known at present, nor its function in HCO3- secretion. ANO6 appears to function as a Ca2+-activated Cl- channel in the intestine that participates in fluid and electrolyte secretion (11), and may have a similar function ion the pancreatic duct. Several other CaCCs are known and are candidates for the ductal CaCC. In pancreatic acinar cells and other serous cells, ANO1 may have a role in HCO3- transport. At physiological [Ca2+]i concentrations ANO1 functions as a Cl- channel. However, at high [Ca2+]i and perhaps at high [Ca2+]i microdomains, ANO1 HCO3- permeability is increased by Ca2+/calmodulin (66, 68), raising the possibility that ANO1 can provide an alternative Cl- and HCO3- conduction in acinar cells (142).
Aquaporins
Although the paracellular pathway is permeable to H2O, H2O flows mostly transcellularly via the water channels aquaporins (AQP) family. This is best illustrated in salivary glands, where knockout of AQP5 markedly reduces salivation (98). Among the 13 AQPs, AQP1 and AQP5 are the major aquaporins in pancreatic duct (17, 73, 74). AQP1 is expressed in the luminal membrane of human acinar and duct cells and is significantly reduced in chronic pancreatitis (156). Moreover, deletion of AQP1 in mice prominently inhibits ductal and pancreatic fluid and HCO3- secretion, due to both reduction in fluid transport and in CFTR expression and activity and thus HCO3- secretion (156). The role of AQP5 in the duct and pancreatic secretion has not been established yet.
Carbonic Anhydrases
A poorly studied topic that deserve more attention is the role of the ductal carbonic anhydrases (CAs) in fluid and electrolyte secretion, in particular with the emerging secretory epithelial diseases due to mutations in CAs. Mutations that affect the action of CA4 cause retinitis pigmentosa (7) and a mutation in CA12 causes salt wasting (32, 106). All transporters involved in fluid and HCO3- secretion are affected by HCO3- concentration at the cellular compartments and microdomains that determine HCO3- availability at plasma membrane inner and outer surfaces. Hydration of CO2 by CAs determines local HCO3- concentration both at the outer and inner plasma membrane surfaces (102). Several CAs are localized in the cytoplasm (such as CA2 and CA7) and several are anchored at the plasma membrane (such as CA4, CA12 and CA14) with the catalytic site at the extracellular surface and regulate HCO3- concentration at the basolateral (CA4 and CA12), or the luminal (CA4) membrane surfaces (33).
CAs localized in the plasma membrane and cytoplasm interact with H+ and HCO3- transporters that mediate ductal fluid and HCO3- secretion and regulate their activity. CA4 interacts with the C terminus of NBCe1-A to increase its activity (6). The C terminus of NBCe1-A and NBCe1-B are conserved and thus it is likely that CA4 regulates NBCe1-B. NBCn1-A recruits the cytoplasmic CA2 to the plasma membrane, where CA2 increases the activity of NBCn1-A (95). CA2 is closely associated with NHE3 and increases NHE3 activity (80). CA2 interacts with a novel site at the C terminus of NHE1 to regulate NHE1 activity (89). CA2 has been reported to interact with the C terminus of slc26a6 to increase its activity. However, the role of other CAs, in particular the plasma membrane anchored CAs, on the activity of the slc26a6 and other SLC26 transporters has not been investigated yet. Finally, CA2 also interacts with AQP1 to increase water flux by AQP1 by an unknown mechanism (158). A particularly interesting CA is the basolateral membrane anchored CA12 with its catalytic site at the extracellular membrane surface. A human mutation in CA12(E143K) is the cause of an autosomal recessive form of salt wasting, which leads to hyponatremia with hyperkalemia, high sweat Cl-, dehydration and failure to thrive. (31, 32, 106). A recent work to understand the cause of the disease established a prominent role for CA12 in ductal fluid and HCO3- secretion. Thus, CA12 increased, while CA12 (E143K) markedly reduced ductal fluid secretion in isolated ducts and in vivo. This could be attributed to a potent stimulation of ductal and topically expressed AE2 by CA12 (53). The E143K mutation is a folding mutation that resulted in retention of CA12(E143K) in the ER (53). How exactly CA12 with an external catalytic site activates AE2 is not obvious. CA12 may clear the extruded HCO3- from the membrane surface to prevent its buildup at the mouth of the AE2. If this can be established, it will be a new mode of regulating HCO3- transporters by CAs.
IV. Regulation and Mechanism of Pancreatic HCO3- Secretion
A. Intracellular Signaling Pathways: cAMP and Ca2+
The cAMP/PKA pathway is central in inducing ductal HCO3- secretion. Secretin is the major hormone that activates the cAMP pathway. VIP also signals to increase cAMP via VIP receptors (VPAC1) (30, 152). At maximal receptor stimulation, the cAMP/PKA pathway can fully activate fluid and HCO3- secretion by activation of the apical CFTR and the basolateral Na+-HCO3- cotransporter, NBCe1-B (165). However, at physiological conditions the cAMP/PKA pathway synergizes with the Ca2+ signaling pathway to activate the secretory process (see below).
Several agonists that act on the pancreatic duct engage the Ca2+ signaling pathway. These include CCK, cholinergic stimuli, P2Rs, and protease-activated receptor 2 (PAR2) receptors (71, 124). When activated, the CCK and muscarinic receptors activate PLCβ to generate IP3 that releases Ca2+ from intracellular stores, mainly the endoplasmic reticulum (ER) and activates the membrane Ca2+ influx channels, Orai and TRPC. P2Rs (96, 110) and PAR2 (8, 107, 109, 111) also act through activation of the Ca2+ signaling pathway. At physiological stimulus intensity, the cAMP and Ca2+ signaling pathways synergize to activate ductal secretion (67). Early studies in vivo already noted the synergistic action of ductal stimuli. Application of secretin at a level observed in the postprandial state only produces modest HCO3- and fluid output (28, 45). Application of CCK and stimulation of M1 and M3 receptors markedly augmented secretin-stimulated pancreatic fluid secretion, although alone CCK and muscarinic stimulation have minimal effect on ductal secretion (86, 167). The molecular mechanism of synergism was resolved with the discovery of regulation of ductal secretion by IRBIT which is discussed below. The cAMP and Ca2+ signaling pathways crosstalk on several additional levels to modulate the activity of each other (67, 121). cAMP/PKA phosphorylates IP3R2 to augment Ca2+ release from the ER (16). Ca2+ influx through the Orai1 channels activates the Ca2+-dependent adenylyl cyclase (AC) AC8 (160). Ca2+ can also activate the CFTR-dependent Cl−/HCO3− exchange activity in CAPAN-1 human pancreatic duct cells (108), which may involve activation by IRBIT.
B. Regulation by IRBIT
Activation of NBCe1-B, slc26a6, and CFTR
IRBIT was isolated as a protein that interacts with the IP3 binding pocket of the receptors (IP3Rs) and it can be dissociated from the IP3Rs by IP3 (25). IRBIT competes with IP3 for binding to the IP3Rs (10) to inhibit Ca2+ release. In fact, the IP3Rs appear to function as IRBIT buffers to prevent IRBIT access to many transporters and targets regulated by IRBIT (87). The C-terminal region of IRBIT family proteins shows ~ 50% homology with the ubiquitous housekeeping enzyme S-adenosyl-l-homocysteine hydrolase (AHCY), with IRBIT having additional N terminal sequence while it lacks the hydrolase activity (9). The main known domains of IRBIT are PP1 and calcineurin binding motif, a PEST domain, a coiled-coil domain, and a PDZ ligand at the end of C terminus (87, 153).
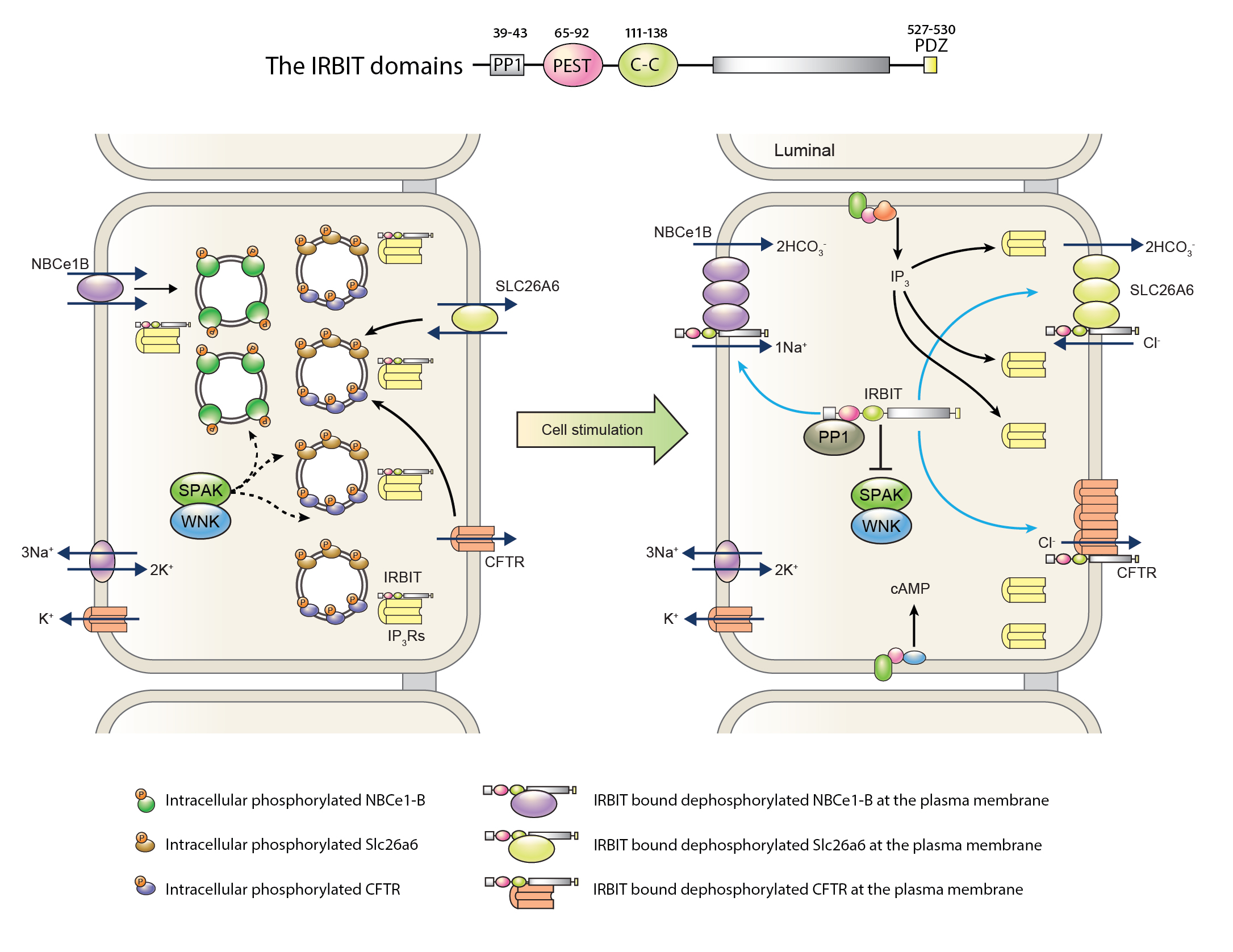
IRBIT plays an important role in pancreatic ductal secretion by regulating multiple transporters and mediating the synergistic action of the cAMP/PKA and Ca2+ signaling pathways (Figure 2). Knockdown of IRBIT in ducts and knockout in mice modestly inhibit fully stimulated pancreatic duct fluid and HCO3- secretion (165), and eliminates the physiological synergistic action of the cAMP/PKA and Ca2+ signaling pathways (122). IRBIT accumulates at the apical pole where IP3Rs are highly expressed, but it can be found all over the cell where IP3Rs are present (88). A search for IRBIT binding proteins identified NBCe1-B as a binding partner, where IRBIT binds to the N terminus autoinhibitory domain of NBCe1-B to activate it by removing the autoinhibition (143). Subsequent detailed studies, in particular with the pancreatic duct revealed that IRBIT at the apical pole potently activates the apical CFTR (163, 165), SLC26A6 (122), and NHE3 (49). At the basal side, IRBIT regulates NBCe1-B (143, 163, 165). IRBIT activates the transporters by multiple mechanisms. First, IRBIT recruits protein phosphatase 1 (PP1) to the transporters to dephosphorylate serine residue 75 in NBCe1-B and yet to be identified residue in CFTR that are phosphorylated by the kinase SPAK. For these phosphorylations SPAK must be activated by phosphorylated by the two kinases WNK1 and/or WNK4 (138). IRBIT also recruits the phosphatase calcineurin to dephosphorylate serine residue 12 that is phosphorylated by CaMKII (153). This enhances the plasma membrane relocation of NBCe1-B, CFTR (163) and slc26a6 (122) from intracellular vesicular pools. At the plasma membrane, IRBIT directly interacts with the transporters to further increase their activity. Moreover, phosphorylation by SPAK and CaMKII and dephosphorylation by the respective phosphatases PP1 and calcineurin determines regulation of NBCe1-B, and likely other IRBIT-regulated transporters, by Cl-in (153). The mechanism by which IRBIT activates the other transporters is not known at this time beyond the need for the PDZ binding motif of IRBIT for assembling the IRBIT-NBCe1-B and IRBIT-CFTR complex (165).
Figure 2. A model for IRBIT associated pathway of pancreatic ductal fluid and HCO3- secretion. Key domains of IRBIT related to HCO3- secretion are illustrated at the top of the figure. In the resting state, IRBIT is bound to IP3Rs, and SPAK phosphorylates NBCe1-B, Slc26a6, and CFTR located at intracellular organelle. When the duct cells are stimulated, IP3 is released and bound to IP3Rs, while IRBIT is disengaged from IP3Rs. PP1 recruited to IRBIT dephosphorylates transporters located at the plasma membrane. IRBIT also binds to the autoinhibitory domain of NBCe1-B to activate it. Increased surface expression of the transporters also aids pancreatic ductal HCO3- secretion. Modified from (121). See text for details.
IRBIT and Synergism
An important action of IRBIT is mediating the synergistic action of the cAMP/PKA and Ca2+ signaling pathways (122) (see Figure 2). Physiological stimulus intensity must be quite weak to prevent cell toxicity that occurs under strong stimulation of all signaling pathways. Indeed, at physiological stimulus intensity the secretory process is activated only by about 5-10% or less of maximal stimulation. Synergism between weakly stimulated signaling is used to generate the maximal response while avoiding cell toxicity and increasing fidelity. IRBIT mediates the synergism between the cAMP/PKA and Ca2+ signaling pathways by translocation between cellular compartments and transporters. At the resting state, IRBIT is sequestered by the high level of IP3Rs at the ductal ER apical pole and is not available for interaction with the transporters. The affinity of the IP3Rs for IRBIT and IP3 is regulated by PKA-mediated phosphorylation of specific IP3Rs serine residues. Phosphorylation of the serine residues increases the affinity for IP3 and at the same time decreases the affinity for IRBIT. Now, a small increase in IP3 evoked by weak stimulation of the Ca2+ signaling pathway is sufficient to dissociate IRBIT from the IP3Rs (122). The released IRBIT can bind to CFTR and slc26a6 first in intracellular vesicles to dephosphorylate them by the IRBIT-recruited PP1 and calcineurin and promote their translocation to the luminal membrane. At the luminal membrane, IRBIT activates the transporters and reduce their inhibition by Cl-in to initiate ductal fluid and HCO3- secretion (122, 153). Of note, the synergistic action of the cAMP/PKA and Ca2+ signaling pathways is eliminated by the knockout of IRBIT (122), highlighting the key role of IRBIT in the synergistic action of the cAMP/PKA and Ca2+ signaling pathways, which is the physiological way that ductal fluid and HCO3- secretion take place.
C. Regulation by [Cl-]i
WNK1 and dynamic regulation of CFTR HCO3- permeability
The WNK proteins consist of four members (WNK1 – WNK4) with a conserved kinase domain that is noted for the unique position of the catalytic lysine residue (105). The discovery that mutations in WNK1 and WNK4 cause hypertension in humans has attracted much attention to these kinases function and regulation (161). The main function of the WNKs is the regulation of Na+, K+, Cl-, HCO3-, and Ca2+ transporters in epithelia and brain (34, 54, 55, 121). The WNKs act either by regulating surface expression of membrane transporters through modulation of their endocytosis or by phosphorylating the transporters and other target proteins directly or indirectly through affecting the effect of other kinases (55). Several functions of WNKs are mediated by phosphorylating and activating the downstream oxidative stress-responsive kinase 1 (OSR1) and SPAK (26). WNK1, WNK3, WNK4, SPAK, and OSR1 are expressed in the pancreatic duct (119, 163) and participate in the regulation of HCO3- transporters and channels (87). Accordingly, knockdown of WNK4 alone or a combined knockdown of WNK1, WNK3 and WNK4 increase pancreatic duct fluid secretion by removing a tonic negative effect on ductal HCO3- transporters (163). However, the role of the WNKs, in particular WNK1, changes at the terminal portion on the duct when [Cl-]i is reduced to below 10 mM. WNK1 and the other WNKs, binds [Cl-]i and their activity is regulated by [Cl-]i (125, 141, 151).
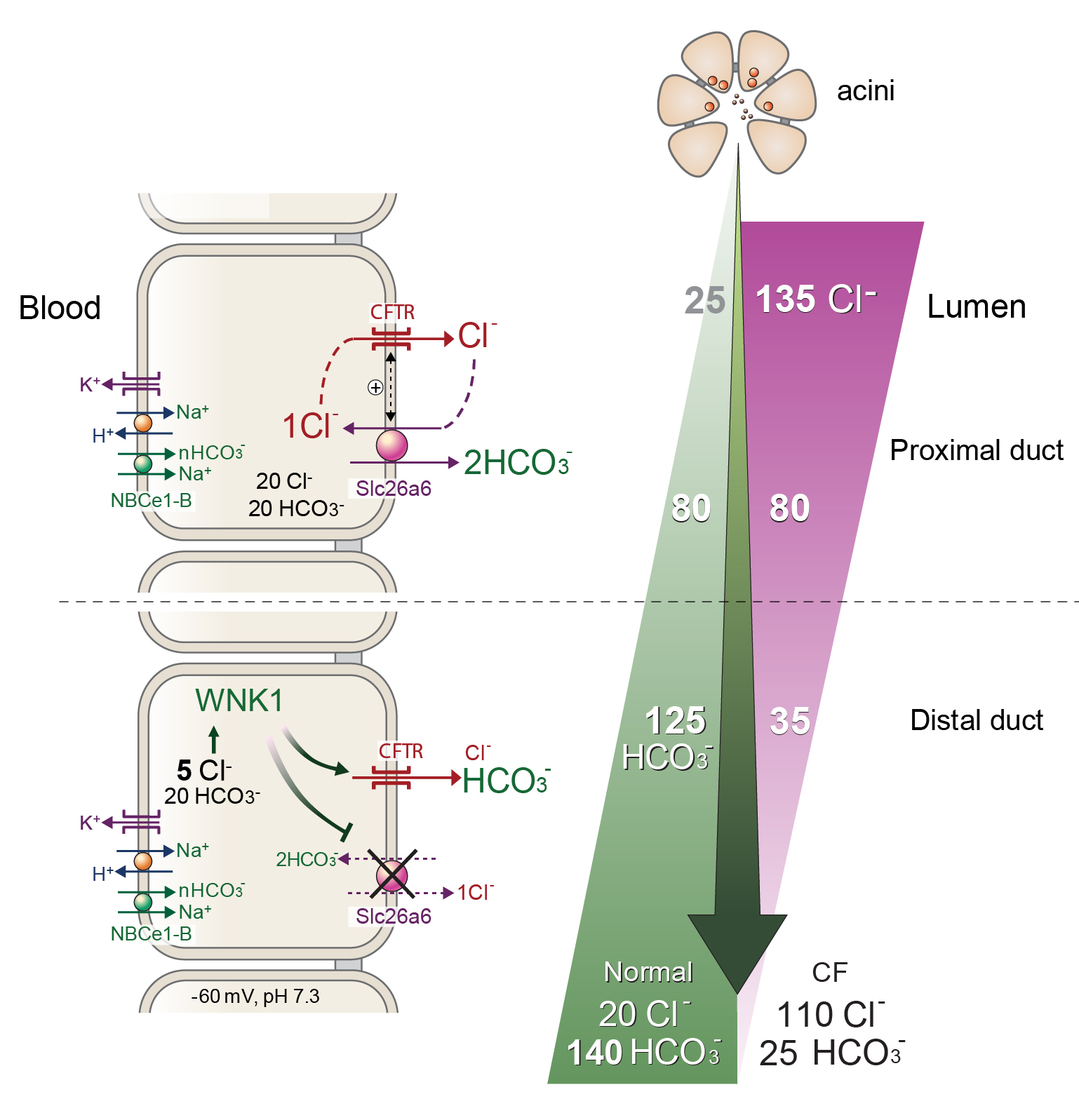
The role of WNK1 in pancreatic HCO3- secretion is illustrated in the left portion of Figure 3. Osmotic stress or low [Cl−]i activates WNK1 (125). Notably, activation of WNK1 by low [Cl−]i greatly increases the HCO3- permeability of CFTR (66, 119). During active pancreatic HCO3- secretion, lower Cl- concentration in the pancreatic juice progressively reduces Cl- absorption. Because of the low basolateral and high luminal Cl- permeability (58, 119), [Cl−]i rapidly decreases in response to the reduction in luminal duct Cl- concentration. At a membrane potential of -60 mV, [Cl−]i is less than 1/10 of luminal Cl- concentration. Indeed, ductal [Cl−]i was estimated to be about 5 mM during cAMP-induced active secretion (58, 119).
Figure 3. A model depicting WNK1-mediated regulation of CFTR in pancreatic ductal function. During active pancreatic HCO3- secretion, Cl- concentration in the pancreatic juice is progressively reduced due to Cl-/HCO3- exchange activities at the apical membrane of duct cells. Because the basolateral membrane of duct cells has poor Cl- permeability but the apical Cl- permeability is very high due to activation of CFTR, [Cl-]i rapidly decreases in response to the reduction in luminal Cl- concentration. Activation of WNK1 by low [Cl-]i increases the PHCO3/PCl of CFTR to over 1.0, which greatly augments HCO3- flux through the CFTR pore. Simultaneously, WNK1/SPAK pathway inhibits Slc26a6 to prevent HCO3- reabsorption. This mechanism enables an increase to as much as 140 mM HCO3- in pancreatic juice. See text for details. Modified from (87).
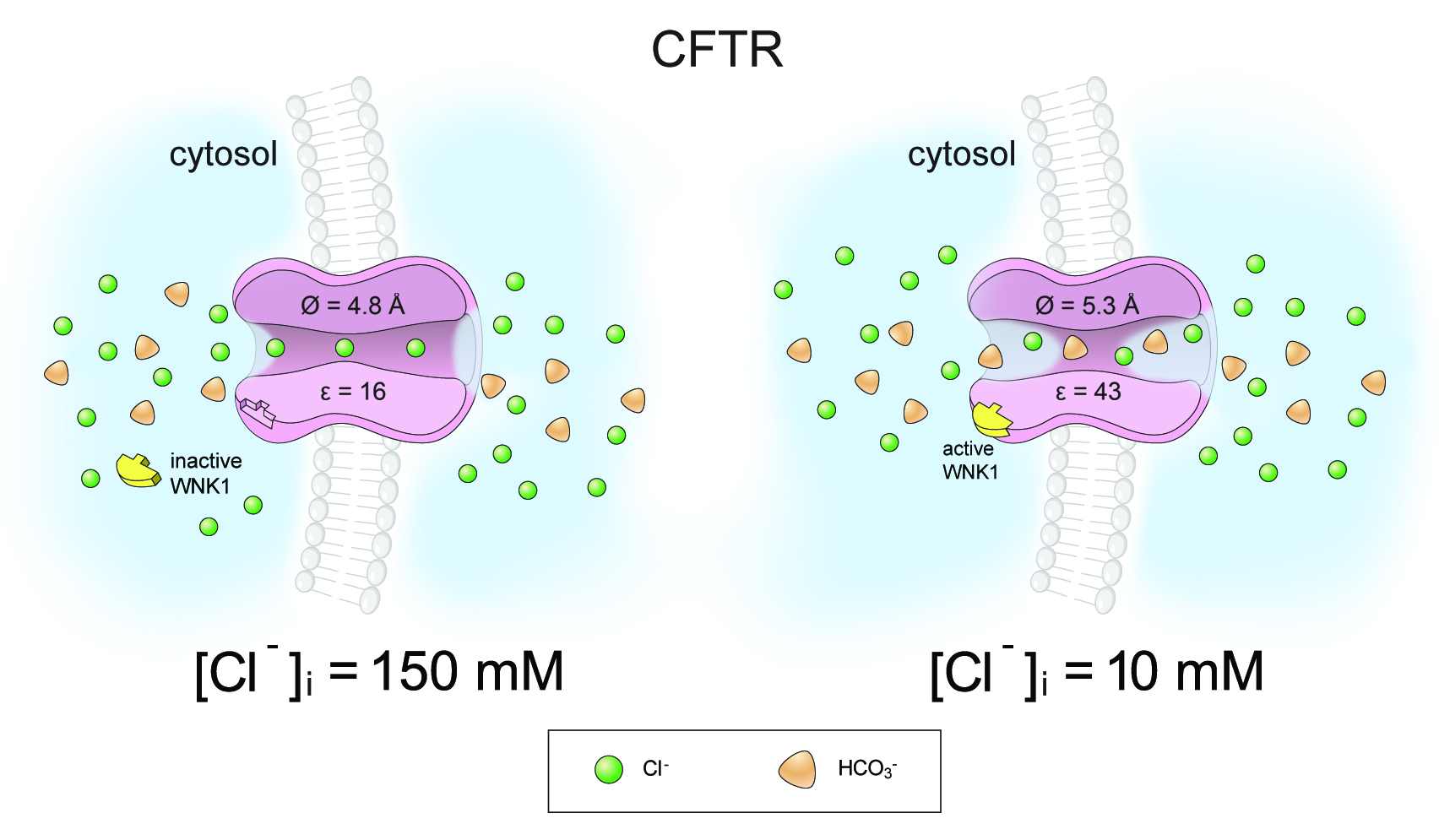
WNK1 modulates the anion selectivity of CFTR by changing its pore size (66). Stimulation by WNK1 increases CFTR pore size from 4.8 Å to 5.3 Å, which facilitates the passage of HCO3-. (4.3 Å, diameter), more than the smaller anion, Cl- (3.7 Å, diameter). Changes in pore size affect the energy barrier of ion dehydration by altering the electric permittivity of the water-filled cavity in the pore. Dielectric constant (relative permittivity, ε) is a unit of electric permittivity, and the dielectric constant of water (εw) is approximately 80 at room temperature. Water molecules in confined geometry like ion channels exhibit a space-dependent reduction in the pore water εw down to 20, due to the restriction of the translational and rotational mobility of water molecules (2). Pore dilation relieves this restriction of water molecule movement and increases εw, which eventually leads to an increase in the overall ε of the anion channel pore. Indeed, the pore dilation induced by WNK1 activation increased the ε of the CFTR pore from 16 to 43 (66). In general, ions pass through the channel after dehydration (at least partial dehydration). Asymmetrically charged ions, such as HCO3-, show lower permeability than the symmetrically charged ions, such as Cl-, due to the high hydration/dehydration energy barrier. The increase in anion channel pore ε greatly alleviates the dehydration penalty of the asymmetrically charged HCO3- and increases PHCO3/PCl (Figure 4). In an initial study, WNK-OSR1 or WNK1-SPAK complex was suggested to increase the CFTR PHCO3/PCl (119). However subsequent study showed that WNK1 alone was sufficient to increase CFTR PHCO3/PCl (70). Molecular dissection of the WNK1 domains revealed that the WNK1 kinase domain is responsible for CFTR PHCO3/PCl selectivity by direct association with CFTR, while the surrounding N-terminal regions mediate the [Cl−]i-sensitivity of WNK1 (70).
Figure 4. WNK1 modulates the anion selectivity of CFTR by changing the pore size. Stimulation by WNK1 increases the pore size of CFTR from 4.8 Å to 5.3 Å and the pore dilation increases the dielectric constant (ε) of the CFTR pore from 16 to 43. The increase in pore size facilitates the passage of the larger anion, HCO3- (4.3 Å, diameter), more than the smaller anion, Cl- (3.7 Å, diameter) by reducing the energy barriers of size-exclusion. More importantly, the dielectric constant increase enhances the HCO3− permeability of CFTR by reducing energy barriers required for ion dehydration of HCO3− (66). See text for details.
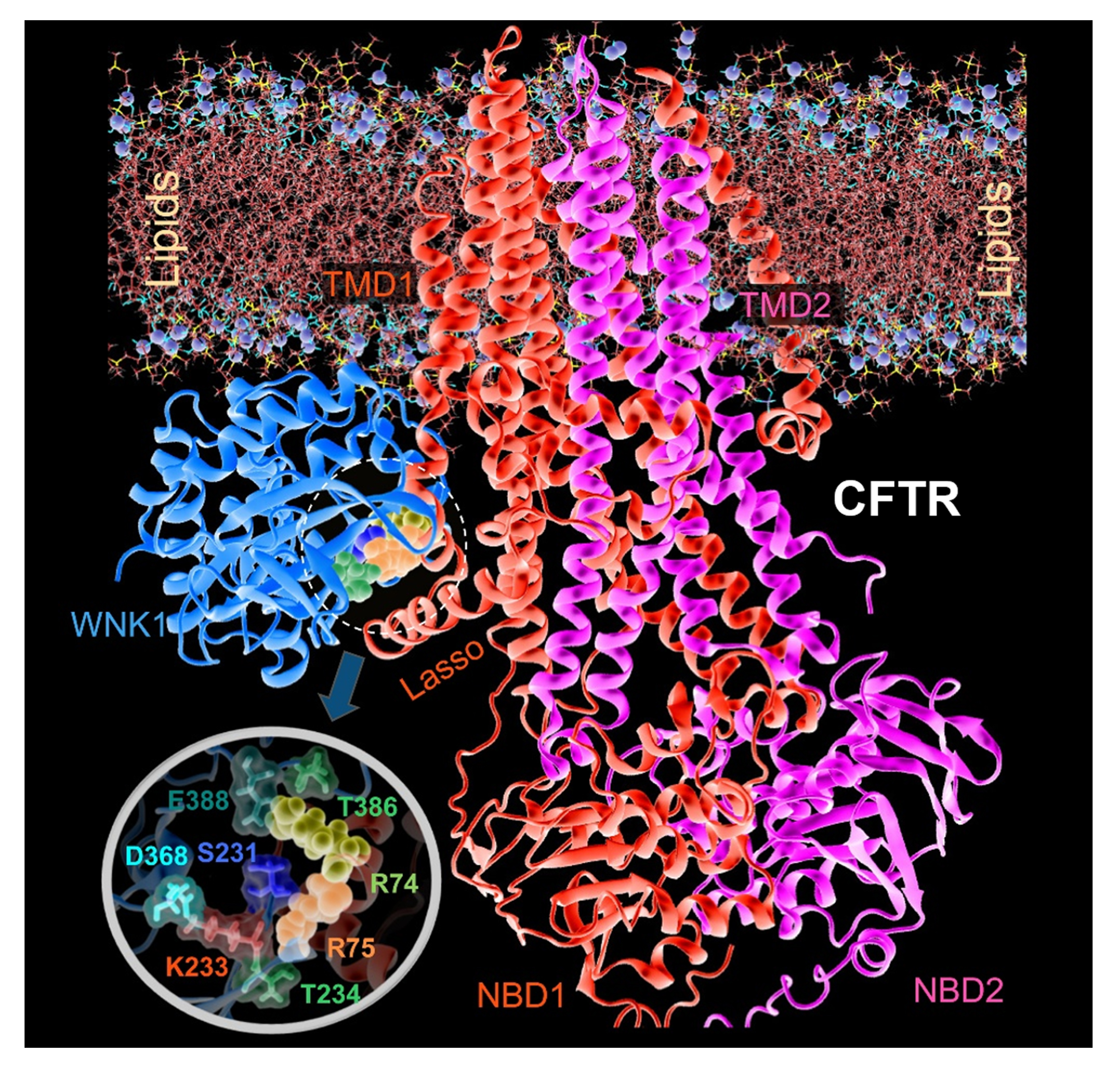
Although the precise WNK1-binding sites on the CFTR are not fully defined, examining pancreatitis-causing CFTR mutations revealed that R74 and R75 located in the first elbow helix region of the CFTR, are involved in the WNK1-CFTR association [39]. A computational protein-protein docking analysis using the Protein Data Bank-deposited structures of human CFTR and the WNK1 kinase domain showed that WNK1 S231–T234 and I384–E388 can potentially bind to an intracellular CFTR region near R74–R75 in the elbow helix 1 (Figure 5). The handle-like elbow helix 1 is located immediately ahead of transmembrane domain 1 and contacts a proximally located lasso motif that has been suggested to play a role in CFTR gating and regulation of the R domain (93). Therefore, it appears that the binding of WNK1 kinase domain to the elbow helix 1 region of CFTR affects the CFTR open structure to facilitate HCO3− permeation (142). Notably, WNK1 not only increases PHCO3/PCl of CFTR HCO3− channel but the HCO3− conductance (GHCO3) and Po in single channel recording (70). Increase in CFTR GHCO3 may also significantly contribute to augmenting HCO3− flux across the apical membrane of pancreatic duct cells.
Figure 5. Structural model for the molecular complex between hCFTR and the WNK1 kinase domain in the presence of a lipid bilayer. The R74 (yellow balls) and R75 (orange balls) residues from hCFTR participate in the binding interface. The figure shows the hCFTR-WNK1 complex predicted by ClusPro, after equilibration in the MD simulation system where it is embedded into the membrane lipids (lines with their phosphorus atoms shown as purple spheres) and solvated by 0.1 M NaCl solution. The snapshot was taken after 100 ns MD simulations. The inset figure shows a close-up view of interfacial interactions. WNK1 residues at the interface include S231, K233, T234, T386, and E388. Modified from (70).
Interestingly, activated WNK1 while increasing CFTR PHCO3/PCl and GHCO3, does not lose the inhibitory effect on SLC26A6 and SLC26A3 (119). When the luminal HCO3- concentration is greater than 140 mM, continuous activation of apical Cl-/HCO3- exchange would reverse to absorb HCO3- from the lumen. This is more of a problem for the 2Cl-/1HCO3- exchange by slc26a3 and less, if at all, for the 1Cl-/2HCO3- slc26a6, especially at membrane potential of -60 mV across the luminal membrane. However, inhibition of the apical Cl-/HCO3- exchangers is required to prevent the reverse mode of Cl-/HCO3- exchange activity if slc26a3 dominates the exchange when ductal [Cl-]i is below 10 mM and ultimately achieves HCO3- concentration above 140 mM in pancreatic juice (146, 148).
Cl-in as a signaling ion, the case for NBCe1-B
[Cl-]i emerged as a signaling ion by regulating several ion transporters and channels. A comprehensive review of Cl- as a bona fide signaling ion can be found in (97). Here we discuss the signaling function of Cl-in with respect to ductal function. By virtue of regulating the function of the WNK kinases [Cl-]i may affect other transporters regulated by these kinases. A significant discovery is that [Cl-]i profoundly regulates the activity of several Na+-HCO3- cotransporters (NBCs) at the [Cl-]i physiological range (138, 153). [Cl-]i regulates the activity of all NBCs tested NBCe1-B, NBCe1-C, and NBCe1-A. The IRBIT-independent activity of NBCe1-B is inhibited by [Cl-]i between 60-140 mM that is outside the physiological range and may function to inhibit NBCe1-B activity under pathological conditions. Most notably, when activated by IRBIT, NBCe1-B activity is reduced by [Cl-]i in the range of 5-20 mM, where at 20 mM [Cl-]i, NBCe1-B activity is reduced to the basal, IRBIT-independent level. Molecular analysis identified two Cl- interacting motifs at the N terminus of NBCe1-B that mediate high and low affinity inhibition by [Cl-]i. Regulation of NBCe1-B is mediated by sites that contain the GXXXP motif. The first site mediates the high [Cl-]i affinity (5-20 mM) regulation of NBCe1-B and the second site mediates the low [Cl-]i affinity (60-140 mM) regulation of NBCe1-B (138). NBCe2-C activity is not regulated by IRBIT and in this case regulation of NBCe1-C is mediated by a single site containing the GXXXP motif and takes place at [Cl-]i between 10-30 mM. Regulation of NBCe1-A by [Cl-]i is mediated by a cryptic Cl- interacting site containing the GXXXP motif. The cryptic NBCe1-A [Cl-]i interacting sites was unmasked by deletion of residues 29-41. Further analysis showed that interaction of Cl-in with the GXXXP sites is regulated by phosphorylation/dephosphorylation events with SPAK and PP1 acting on serine 65 to affect Cl- sensing by the 32GXXXP36 site, while CaMKII and calcineurin acting on serine 12 to affect Cl- sensing by the 194GXXXP198 site (153). Other phosphorylation sites affecting NBCe1-B activity are Ser232, Ser233, and Ser235 with the phosphorylation status of Ser232, Ser233, and Ser235 is regulated by IRBIT to determine whether NBCe1 transporters are in active or inactive conformations (153).
Hence, cells have a [Cl-]i sensing mechanism that plays an important role in the regulation of Na+ and HCO3- transporters that mediate the critical step of HCO3- influx in the process of ductal fluid and HCO3- secretion. At [Cl-]i of up to 20 mM, CFTR functions mostly as a Cl- channel and slc26a6 mediates most ductal HCO3- secretion. As [Cl-]i is reduced below 20 mM and additional HCO3- secretion takes place in the face of unfavorable Cl- and HCO3- gradients across the apical membrane, there is an increased demand for HCO3- entry across the basolateral membrane. Pancreatic duct cells achieve this by [Cl-]i-mediated regulation of NBCe1-B and CFTR, at which NBCe1-B activity and CFTR HCO3- permeability gradually increase as [Cl-]i is reduced towards 5 mM.
D. A Model for Pancreatic HCO3- Secretion
Electrogenic HCO3− transporters can generate higher gradients of HCO3− than electroneutral transporters when the electro-repulsive force generated by the negative membrane potential is coupled to the basolateral uptake of HCO3- by NBCe1-B and the luminal efflux of HCO3− through SLC26A6 and CFTR. The electrogenic SLC26A6 exchanger with the stoichiometry of 1Cl-: 2HCO3−,can achieve luminal HCO3− concentration of up to about 120 mM at apical membrane potential of -60 mV (148). To drive luminal HCO3- concentration to 140 mM, the physiologic HCO3- concentrations in pancreatic juice, another mechanism is needed (148). Such a mechanism should be Cl- independent, since significant fraction of pancreatic HCO3− secretion is retained in the absence of luminal Cl- (59, 61). The WNK1 activated CFTR satisfy these requirements. At HCO3-in in stimulated duct cells above 25 mM and membrane potential of -60 mV, CFTR mediates HCO3- efflux even at luminal HCO3- concentrations of above 140 mM. Transport by NBCe1-B and slc26a6 results in osmotic transport of HCO3- that is essential for the transcellular water transport by the duct. The transcellular basal to luminal electrogenic HCO3- transport by both slc26a6 and CFTR generates a lumen-negative electrical potential that results in paracellular Na+ secretion. Water flows down the osmotic gradient generated by the Na+ and HCO3- fluxes via paracellular and transcellular (aquaporins) pathways (Figure 3). Overall, these processes generate an efficient mechanism for HCO3-–driven ductal fluid secretion to generate the volume and HCO3- content of the pancreatic juice.
V. Conclusions
The mechanism by which the human pancreatic duct secretes nearly isotonic HCO3- solution has long been an enigmatic question for both physiologists and clinicians (86, 148). When Bayliss and Starling first noticed that the exocrine pancreas secretes alkaline fluid, they assumed that carbonate is responsible for the strong alkalinity of the pancreatic juice (13). Later, with better understanding of the carbonate/HCO3-/CO2 buffer system (51), it became clear that the exocrine pancreas secretes fluid in which the dominant anion is HCO3-, and HCO3− secretion is coupled to fluid secretion (19, 28, 46). Current understanding indicates that activation of three key transporters, the basolateral NBCe1-B (and likely AE2), and the luminal SLC26A6 and CFTR, and their synergistic regulation by the cAMP and Ca2+ signaling pathways through IRBIT and WNK1 perform vectorial pancreatic HCO3− secretion that drives fluid secretion. NBCe1-B, with a 1 Na+/2 HCO3- stoichiometry, is the main HCO3- concentrating transporter in the basolateral membrane, and can achieve the necessary HCO3- influx (1, 60, 172). Basolateral AE2 activity is also required to support ductal HCO3- fluid and HCO3- secretion probably by controlling cytoplasmic and near membrane pHin, although the exact role of AE2 is not known at present. The electrogenic SLC26A6, with a 1 Cl-/2 HCO3- stoichiometry is the major apical Cl-/HCO3- exchanger, which mediates most HCO3- efflux in the early step of pancreatic HCO3− secretion (76, 94). Activated WNK1 increases HCO3- permeability and conductance of CFTR, allowing further apical HCO3- efflux and setting the pancreatic juice HCO3- concentrations above 140 mM (119). Ductal fluid and HCO3- secretion is essential for the function of the pancreas and is severely altered in all form of pancreatitis (50, 168). Our understanding of the mechanism of pancreatic fluid and HCO3- secretion will continue to improve as our knowledge of existing pathways increases and new mechanisms are identified and delineated, to provide a better scientific basis for therapeutic approaches to treat diseases like cystic fibrosis and acute and chronic pancreatitis.
VI. Acknowledgments
We thank Dong-Su Jang for assistance in illustrations. This work was supported by grant, 2013R1A3A2042197, from the National Research Foundation, the Ministry of Science, ICT & Future Planning, Republic of Korea and by Intramural NIH/NIDCR grant DE000735-010.
VII. References
- Abuladze N, Lee I, Newman D, Hwang J, Boorer K, Pushkin A, and Kurtz I. Molecular cloning, chromosomal localization, tissue distribution, and functional expression of the human pancreatic sodium bicarbonate cotransporter. J Biol Chem 273: 17689-17695, 1998. PMID: 9651366.
- Aguilella-Arzo M, Andrio A, Aguilella VM, and Alcaraz A. Dielectric saturation of water in a membrane protein channel. Phys Chem Chem Phys 11: 358-365, 2009. PMID: 19088992.
- Ahn W, Kim KH, Lee JA, Kim JY, Choi JY, Moe OW, Milgram SL, Muallem S, and Lee MG. Regulatory interaction between the cystic fibrosis transmembrane conductance regulator and HCO3- salvage mechanisms in model systems and the mouse pancreatic duct. J Biol Chem 276: 17236-17243, 2001. PMID: 11278980.
- Ahuja M, Chung WY, Lin WY, McNally BA, and Muallem S. Ca(2+) Signaling in Exocrine Cells. Cold Spring Harb Perspect Biol 12: 2020. PMID: 31636079.
- Almassy J, Diszhazi G, Skaliczki M, Marton I, Magyar ZE, Nanasi PP, and Yule DI. Expression of BK channels and Na(+)-K(+) pumps in the apical membrane of lacrimal acinar cells suggests a new molecular mechanism for primary tear-secretion. Ocul Surf 17: 272-277, 2019. PMID: 30685438.
- Alvarez BV, Loiselle FB, Supuran CT, Schwartz GJ, and Casey JR. Direct extracellular interaction between carbonic anhydrase IV and the human NBC1 sodium/bicarbonate co-transporter. Biochemistry 42: 12321-12329, 2003. PMID: 14567693.
- Alvarez BV, Vithana EN, Yang Z, Koh AH, Yeung K, Yong V, Shandro HJ, Chen Y, Kolatkar P, Palasingam P, Zhang K, Aung T, and Casey JR. Identification and characterization of a novel mutation in the carbonic anhydrase IV gene that causes retinitis pigmentosa. Invest Ophthalmol Vis Sci 48: 3459-3468, 2007. PMID: 17652713.
- Alvarez C, Regan JP, Merianos D, and Bass BL. Protease-activated receptor-2 regulates bicarbonate secretion by pancreatic duct cells in vitro. Surgery 136: 669-676, 2004. PMID: 15349117.
- Ando H, Kawaai K, and Mikoshiba K. IRBIT: a regulator of ion channels and ion transporters. Biochim Biophys Acta 1843: 2195-2204, 2014. PMID: 24518248.
- Ando H, Mizutani A, Matsu-ura T, and Mikoshiba K. IRBIT, a novel inositol 1,4,5-trisphosphate (IP3) receptor-binding protein, is released from the IP3 receptor upon IP3 binding to the receptor. J Biol Chem 278: 10602-10612, 2003. PMID: 112525476.
- Aoun J, Hayashi M, Sheikh IA, Sarkar P, Saha T, Ghosh P, Bhowmick R, Ghosh D, Chatterjee T, Chakrabarti P, Chakrabarti MK, and Hoque KM. Anoctamin 6 Contributes to Cl- Secretion in Accessory Cholera Enterotoxin (Ace)-stimulated Diarrhea: AN ESSENTIAL ROLE FOR PHOSPHATIDYLINOSITOL 4,5-BISPHOSPHATE (PIP2) SIGNALING IN CHOLERA. J Biol Chem 291: 26816-26836, 2016. PMID: 27799301.
- Argent BE, and Case RM. Pancreatic ducts. Cellular mechanism and control of bicarbonate secretion. Physiology of the gastrointestinal tract 1473-1497, 1994.
- Bayliss WM, and Starling EH. The mechanism of pancreatic secretion. J Physiol 28: 325-353, 1902. PMID: 16992627.
- Boron WF, Chen L, and Parker MD. Modular structure of sodium-coupled bicarbonate transporters. J Exp Biol 212: 1697-1706, 2009. PMID: 19448079.
- Brooks AM, and Grossman MI. Postprandial pH and neutralizing capacity of the proximal duodenum in dogs. Gastroenterology 59: 85-89, 1970. PMID: 5426993.
- Bruce JI, Shuttleworth TJ, Giovannucci DR, and Yule DI. Phosphorylation of inositol 1,4,5-trisphosphate receptors in parotid acinar cells. A mechanism for the synergistic effects of cAMP on Ca2+ signaling. J Biol Chem 277: 1340-1348, 2002. PMID: 11694504.
- Burghardt B, Elkaer ML, Kwon TH, Racz GZ, Varga G, Steward MC, and Nielsen S. Distribution of aquaporin water channels AQP1 and AQP5 in the ductal system of the human pancreas. Gut 52: 1008-1016, 2003. PMID: 12801959.
- Caputo A, Caci E, Ferrera L, Pedemonte N, Barsanti C, Sondo E, Pfeffer U, Ravazzolo R, Zegarra-Moran O, and Galietta LJ. TMEM16A, a membrane protein associated with calcium-dependent chloride channel activity. Science 322: 590-594, 2008. PMID: 18772398.
- Case RM, Harper AA, and Scratcherd T. The secretion of electrolytes and enzymes by the pancreas of the anaesthetized cat. J Physiol 201: 335-348, 1969. PMID: 5780548.
- Catalan MA, Nakamoto T, Gonzalez-Begne M, Camden JM, Wall SM, Clarke LL, and Melvin JE. Cftr and ENaC ion channels mediate NaCl absorption in the mouse submandibular gland. J Physiol 588: 713-724, 2010. PMID: 20026617.
- Chandra R, and Liddle RA. Regulation of Pancreatic Secretion. The Pancreapedia: Exocrine Pancreas Knowledge Base 2015. DOI: 10.3998/panc.2015.38.
- Chey WY, Lee YH, Hendricks JG, Rhodes RA, and Tai HH. Plasma secretin concentrations in fasting and postprandial state in man. Am J Dig Dis 23: 981-988, 1978. PMID: 31087.
- Choi JY, Muallem D, Kiselyov K, Lee MG, Thomas PJ, and Muallem S. Aberrant CFTR-dependent HCO3- transport in mutations associated with cystic fibrosis. Nature 410: 94-97, 2001. PMID: 11242048.
- Deeley RG, Westlake C, and Cole SP. Transmembrane transport of endo- and xenobiotics by mammalian ATP-binding cassette multidrug resistance proteins. Physiol Rev 86: 849-899, 2006. PMID: 16816140.
- Dekker JW, Budhia S, Angel NZ, Cooper BJ, Clark GJ, Hart DN, and Kato M. Identification of an S-adenosylhomocysteine hydrolase-like transcript induced during dendritic cell differentiation. Immunogenetics 53: 993-1001, 2002. PMID: 11904675.
- Delpire E, and Gagnon KB. SPAK and OSR1: STE20 kinases involved in the regulation of ion homoeostasis and volume control in mammalian cells. Biochem J 409: 321-331, 2008. PMID: 18092945.
- Domschke S, Domschke W, Rosch W, Konturek SJ, Sprugel W, Mitznegg P, Wunsch E, and Demling L. Inhibition by somatostatin of secretin-stimulated pancreatic secretion in man: a study with pure pancreatic juice. Scand J Gastroenterol 12: 59-63, 1977. PMID: 189382.
- Domschke S, Domschke W, Rosch W, Konturek SJ, Wunsch E, and Demling L. Bicarbonate and cyclic AMP content of pure human pancreatic juice in response to graded doses of synthetic secretin. Gastroenterology 70: 533-536, 1976. PMID: 176080.
- Dorwart MR, Shcheynikov N, Yang D, and Muallem S. The solute carrier 26 family of proteins in epithelial ion transport. Physiology (Bethesda) 23: 104-114, 2008. PMID: 18400693.
- Evans RL, Perrott MN, Lau KR, and Case RM. Elevation of intracellular cAMP by noradrenaline and vasoactive intestinal peptide in striated ducts isolated from the rabbit mandibular salivary gland. Arch Oral Biol 41: 689-694, 1996. PMID: 9015570.
- Feinstein Y, Yerushalmi B, Loewenthal N, Alkrinawi S, Birk OS, Parvari R, and Hershkovitz E. Natural history and clinical manifestations of hyponatremia and hyperchlorhidrosis due to carbonic anhydrase XII deficiency. Horm Res Paediatr 81: 336-342, 2014. PMID: 24714577.
- Feldshtein M, Elkrinawi S, Yerushalmi B, Marcus B, Vullo D, Romi H, Ofir R, Landau D, Sivan S, Supuran CT, and Birk OS. Hyperchlorhidrosis caused by homozygous mutation in CA12, encoding carbonic anhydrase XII. Am J Hum Genet 87: 713-720, 2010. PMID: 21035102.
- Frost SC. Physiological functions of the alpha class of carbonic anhydrases. Subcell Biochem 75: 9-30, 2014. PMID: 24146372.
- Furusho T, Uchida S, and Sohara E. The WNK signaling pathway and salt-sensitive hypertension. Hypertens Res 43: 733-743, 2020. PMID: 32286498.
- Gagnon KB, and Delpire E. Molecular physiology of SPAK and OSR1: two Ste20-related protein kinases regulating ion transport. Physiol Rev 92: 1577-1617, 2012. PMID: 23073627.
- Gautam D, Han SJ, Heard TS, Cui Y, Miller G, Bloodworth L, and Wess J. Cholinergic stimulation of amylase secretion from pancreatic acinar cells studied with muscarinic acetylcholine receptor mutant mice. J Pharmacol Exp Ther 313: 995-1002, 2005. PMID: 15764735.
- Gerolami A, Marteau C, Matteo A, Sahel J, Portugal H, Pauli AM, Pastor J, and Sarles H. Calcium carbonate saturation in human pancreatic juice: possible role of ductal H+ secretion. Gastroenterology 96: 881-884, 1989. PMID: 2914648.
- Gray MA, Greenwell JR, Garton AJ, and Argent BE. Regulation of maxi-K+ channels on pancreatic duct cells by cyclic AMP-dependent phosphorylation. J Membr Biol 115: 203-215, 1990. PMID: 1695685.
- Gray MA, Harris A, Coleman L, Greenwell JR, and Argent BE. Two types of chloride channel on duct cells cultured from human fetal pancreas. Am J Physiol 257: C240-251, 1989. PMID: 2475028.
- Gray MA, Winpenny JP, Porteous DJ, Dorin JR, and Argent BE. CFTR and calcium-activated chloride currents in pancreatic duct cells of a transgenic CF mouse. Am J Physiol 266: C213-221, 1994. PMID: 7508188.
- Gross E, Fedotoff O, Pushkin A, Abuladze N, Newman D, and Kurtz I. Phosphorylation-induced modulation of pNBC1 function: distinct roles for the amino- and carboxy-termini. J Physiol 549: 673-682, 2003. PMID: 12730338.
- Gross E, Hawkins K, Abuladze N, Pushkin A, Cotton CU, Hopfer U, and Kurtz I. The stoichiometry of the electrogenic sodium bicarbonate cotransporter NBC1 is cell-type dependent. J Physiol 531: 597-603, 2001. PMID: 11251043.
- Grundy D, Hutson D, and Scratcherd T. The response of the pancreas of the anaesthetized cat to secretin before, during and after reversible vagal blockade. J Physiol 342: 517-526, 1983. PMID: 6631748.
- Guggino WB. The cystic fibrosis transmembrane regulator forms macromolecular complexes with PDZ domain scaffold proteins. Proc Am Thorac Soc 1: 28-32, 2004. PMID: 16113408.
- Gyr K, Beglinger C, Fried M, Grotzinger U, Kayasseh L, Stalder GA, and Girard J. Plasma secretin and pancreatic response to various stimulants including a meal. Am J Physiol 246: G535-542, 1984. PMID: 6720952.
- Hart WM, and Thomas JE. Bicarbonate and chloride of pancreatic juice secreated in response to various stimuli. Gastroenterology 4: 1945.
- Hatefi Y, and Hanstein WG. Solubilization of particulate proteins and nonelectrolytes by chaotropic agents. Proc Natl Acad Sci U S A 62: 1129-1136, 1969. PMID: 5256411.
- He P, Klein J, and Yun CC. Activation of Na+/H+ exchanger NHE3 by angiotensin II is mediated by inositol 1,4,5-triphosphate (IP3) receptor-binding protein released with IP3 (IRBIT) and Ca2+/calmodulin-dependent protein kinase II. J Biol Chem 285: 27869-27878, 2010. PMID: 20584908.
- He P, Zhang H, and Yun CC. IRBIT, inositol 1,4,5-triphosphate (IP3) receptor-binding protein released with IP3, binds Na+/H+ exchanger NHE3 and activates NHE3 activity in response to calcium. J Biol Chem 283: 33544-33553, 2008. PMID: 18829453.
- Hegyi P, Wilschanski M, Muallem S, Lukacs GL, Sahin-Toth M, Uc A, Gray MA, Rakonczay Z, Jr., and Maleth J. CFTR: A New Horizon in the Pathomechanism and Treatment of Pancreatitis. Rev Physiol Biochem Pharmacol 170: 37-66, 2016. PMID: 26856995.
- Henderson LJ. The Regulation of Neutrality in the Animal Body. Science 37: 389-395, 1913. PMID: 17795147.
- Holst JJ, Fahrenkrug J, Knuhtsen S, Jensen SL, Poulsen SS, and Nielsen OV. Vasoactive intestinal polypeptide (VIP) in the pig pancreas: role of VIPergic nerves in control of fluid and bicarbonate secretion. Regul Pept 8: 245-259, 1984. PMID: 6379759.
- Hong JH, Muhammad E, Zheng C, Hershkovitz E, Alkrinawi S, Loewenthal N, Parvari R, and Muallem S. Essential role of carbonic anhydrase XII in secretory gland fluid and HCO3 (-) secretion revealed by disease causing human mutation. J Physiol 593: 5299-5312, 2015. PMID: 26486891.
- Huang CL, Cha SK, Wang HR, Xie J, and Cobb MH. WNKs: protein kinases with a unique kinase domain. Exp Mol Med 39: 565-573, 2007. PMID: 18059132.
- Huang CL, Yang SS, and Lin SH. Mechanism of regulation of renal ion transport by WNK kinases. Curr Opin Nephrol Hypertens 17: 519-525, 2008. PMID: 18695394.
- Hug MJ, Tamada T, and Bridges RJ. CFTR and bicarbonate secretion by [correction of to] epithelial cells. News Physiol Sci 18: 38-42, 2003. PMID: 12531931.
- Ishiguro H, Naruse S, Kitagawa M, Hayakawa T, Case RM, and Steward MC. Luminal ATP stimulates fluid and HCO3- secretion in guinea-pig pancreatic duct. J Physiol 519 Pt 2: 551-558, 1999. PMID: 10457070.
- Ishiguro H, Naruse S, Kitagawa M, Mabuchi T, Kondo T, Hayakawa T, Case RM, and Steward MC. Chloride transport in microperfused interlobular ducts isolated from guinea-pig pancreas. J Physiol 539: 175-189, 2002. PMID: 11850511.
- Ishiguro H, Naruse S, Steward MC, Kitagawa M, Ko SB, Hayakawa T, and Case RM. Fluid secretion in interlobular ducts isolated from guinea-pig pancreas. J Physiol 511 ( Pt 2): 407-422, 1998. PMID: 9706019.
- Ishiguro H, Steward MC, Lindsay AR, and Case RM. Accumulation of intracellular HCO3- by Na(+)-HCO3- cotransport in interlobular ducts from guinea-pig pancreas. J Physiol 495 ( Pt 1): 169-178, 1996. PMID: 8866360.
- Ishiguro H, Steward MC, Naruse S, Ko SB, Goto H, Case RM, Kondo T, and Yamamoto A. CFTR functions as a bicarbonate channel in pancreatic duct cells. J Gen Physiol 133: 315-326, 2009. PMID: 19204187.
- Ishiguro H, Steward MC, Wilson RW, and Case RM. Bicarbonate secretion in interlobular ducts from guinea-pig pancreas. J Physiol 495 ( Pt 1): 179-191, 1996. PMID: 8866361.
- Jha A, Chung WY, Vachel L, Maleth J, Lake S, Zhang G, Ahuja M, and Muallem S. Anoctamin 8 tethers endoplasmic reticulum and plasma membrane for assembly of Ca(2+) signaling complexes at the ER/PM compartment. EMBO J 38: 2019. PMID: 31061173.
- Jin C, Naruse S, Kitagawa M, Ishiguro H, Nakajima M, Mizuno N, Ko SB, and Hayakawa T. The effect of calcitonin gene-related peptide on pancreatic blood flow and secretion in conscious dogs. Regul Pept 99: 9-15, 2001. PMID: 11257309.
- Johansen PG, Anderson CM, and Hadorn B. Cystic fibrosis of the pancreas. A generalised disturbance of water and electrolyte movement in exocrine tissues. Lancet 1: 455-460, 1968. PMID: 4170642.
- Jun I, Cheng MH, Sim E, Jung J, Suh BL, Kim Y, Son H, Park K, Kim CH, Yoon JH, Whitcomb DC, Bahar I, and Lee MG. Pore dilatation increases the bicarbonate permeability of CFTR, ANO1 and glycine receptor anion channels. J Physiol 594: 2929-2955, 2016. PMID: 26663196.
- Jung J, and Lee MG. Role of calcium signaling in epithelial bicarbonate secretion. Cell Calcium 55: 376-384, 2014. PMID: 24598807.
- Jung J, Nam JH, Park HW, Oh U, Yoon JH, and Lee MG. Dynamic modulation of ANO1/TMEM16A HCO3(-) permeability by Ca2+/calmodulin. Proc Natl Acad Sci U S A 110: 360-365, 2013. PMID: 23248295.
- Kerem B, Rommens JM, Buchanan JA, Markiewicz D, Cox TK, Chakravarti A, Buchwald M, and Tsui LC. Identification of the cystic fibrosis gene: genetic analysis. Science 245: 1073-1080, 1989. PMID: 2570460.
- Kim Y, Jun I, Shin DH, Yoon JG, Piao H, Jung J, Park HW, Cheng MH, Bahar I, Whitcomb DC, and Lee MG. Regulation of CFTR Bicarbonate Channel Activity by WNK1: Implications for Pancreatitis and CFTR-Related Disorders. Cell Mol Gastroenterol Hepatol 9: 79-103, 2020. PMID: 31561038.
- Kiselyov K, Wang X, Shin DM, Zang W, and Muallem S. Calcium signaling complexes in microdomains of polarized secretory cells. Cell Calcium 40: 451-459, 2006. PMID: 17034849.
- Knauf F, Yang CL, Thomson RB, Mentone SA, Giebisch G, and Aronson PS. Identification of a chloride-formate exchanger expressed on the brush border membrane of renal proximal tubule cells. Proc Natl Acad Sci U S A 98: 9425-9430, 2001. PMID: 11459928.
- Ko SB, Mizuno N, Yatabe Y, Yoshikawa T, Ishiguro H, Yamamoto A, Azuma S, Naruse S, Yamao K, Muallem S, and Goto H. Corticosteroids correct aberrant CFTR localization in the duct and regenerate acinar cells in autoimmune pancreatitis. Gastroenterology 138: 1988-1996, 2010. PMID: 20080093.
- Ko SB, Naruse S, Kitagawa M, Ishiguro H, Furuya S, Mizuno N, Wang Y, Yoshikawa T, Suzuki A, Shimano S, and Hayakawa T. Aquaporins in rat pancreatic interlobular ducts. Am J Physiol Gastrointest Liver Physiol 282: G324-331, 2002. PMID: 11804854.
- Ko SB, Shcheynikov N, Choi JY, Luo X, Ishibashi K, Thomas PJ, Kim JY, Kim KH, Lee MG, Naruse S, and Muallem S. A molecular mechanism for aberrant CFTR-dependent HCO(3)(-) transport in cystic fibrosis. EMBO J 21: 5662-5672, 2002. PMID: 12411484.
- Ko SB, Zeng W, Dorwart MR, Luo X, Kim KH, Millen L, Goto H, Naruse S, Soyombo A, Thomas PJ, and Muallem S. Gating of CFTR by the STAS domain of SLC26 transporters. Nat Cell Biol 6: 343-350, 2004. PMID: 15048129.
- Kohler H, Nustede R, Barthel M, and Schafmayer A. Exocrine pancreatic function in dogs with denervated pancreas. Pancreas 2: 676-683, 1987. PMID: 3325985.
- Konturek SJ, Zabielski R, Konturek JW, and Czarnecki J. Neuroendocrinology of the pancreas; role of brain-gut axis in pancreatic secretion. Eur J Pharmacol 481: 1-14, 2003. PMID: 14637169.
- Kowal JM, Yegutkin GG, and Novak I. ATP release, generation and hydrolysis in exocrine pancreatic duct cells. Purinergic Signal 11: 533-550, 2015. PMID: 26431833.
- Krishnan D, Liu L, Wiebe SA, Casey JR, Cordat E, and Alexander RT. Carbonic anhydrase II binds to and increases the activity of the epithelial sodium-proton exchanger, NHE3. Am J Physiol Renal Physiol 309: F383-392, 2015. PMID: 26041446.
- Kunzelmann K, Schreiber R, and Hadorn HB. Bicarbonate in cystic fibrosis. J Cyst Fibros 16: 653-662, 2017. PMID: 28732801.
- Kusiak AA, Szopa MD, Jakubowska MA, and Ferdek PE. Signaling in the Physiology and Pathophysiology of Pancreatic Stellate Cells - a Brief Review of Recent Advances. Front Physiol 11: 78, 2020. PMID: 32116785.
- LaRusch J, Jung J, General IJ, Lewis MD, Park HW, Brand RE, Gelrud A, Anderson MA, Banks PA, Conwell D, Lawrence C, Romagnuolo J, Baillie J, Alkaade S, Cote G, Gardner TB, Amann ST, Slivka A, Sandhu B, Aloe A, Kienholz ML, Yadav D, Barmada MM, Bahar I, Lee MG, Whitcomb DC, and North American Pancreatitis Study G. Mechanisms of CFTR functional variants that impair regulated bicarbonate permeation and increase risk for pancreatitis but not for cystic fibrosis. PLoS Genet 10: e1004376, 2014. PMID: 25033378.
- Lee JH, Richter W, Namkung W, Kim KH, Kim E, Conti M, and Lee MG. Dynamic regulation of cystic fibrosis transmembrane conductance regulator by competitive interactions of molecular adaptors. J Biol Chem 282: 10414-10422, 2007. PMID: 17244609.
- Lee MG, Ahn W, Choi JY, Luo X, Seo JT, Schultheis PJ, Shull GE, Kim KH, and Muallem S. Na(+)-dependent transporters mediate HCO(3)(-) salvage across the luminal membrane of the main pancreatic duct. J Clin Invest 105: 1651-1658, 2000. PMID: 10841524.
- Lee MG, and Muallem S. Physiology of duct cell secretion. In: Pancreas: An Integrated Textbook of Basic Science, Medicine, and Surgery, edited by Beger H BM, Kozarek R, Lerch M, Neoptolemos J, Warshaw A, Whitcomb D, Shiratori K. . Oxford, U.K: Blackwell Publishing, 2008, p. 78-90.
- Lee MG, Ohana E, Park HW, Yang D, and Muallem S. Molecular mechanism of pancreatic and salivary gland fluid and HCO3 secretion. Physiol Rev 92: 39-74, 2012. PMID: 22298651.
- Lee MG, Xu X, Zeng W, Diaz J, Wojcikiewicz RJ, Kuo TH, Wuytack F, Racymaekers L, and Muallem S. Polarized expression of Ca2+ channels in pancreatic and salivary gland cells. Correlation with initiation and propagation of [Ca2+]i waves. J Biol Chem 272: 15765-15770, 1997. PMID: 9188472.
- Li X, Liu Y, Alvarez BV, Casey JR, and Fliegel L. A novel carbonic anhydrase II binding site regulates NHE1 activity. Biochemistry 45: 2414-2424, 2006. PMID: 16475831.
- Lifson N, Kramlinger KG, Mayrand RR, and Lender EJ. Blood flow to the rabbit pancreas with special reference to the islets of Langerhans. Gastroenterology 79: 466-473, 1980. PMID: 7000613.
- Lin WY, and Muallem S. No Zoom Required: Meeting at the beta-Intercalated Cells. J Am Soc Nephrol 31: 1655-1657, 2020. PMID: 32716314.
- Linsdell P, Tabcharani JA, Rommens JM, Hou YX, Chang XB, Tsui LC, Riordan JR, and Hanrahan JW. Permeability of wild-type and mutant cystic fibrosis transmembrane conductance regulator chloride channels to polyatomic anions. J Gen Physiol 110: 355-364, 1997. PMID: 9379168.
- Liu F, Zhang Z, Csanady L, Gadsby DC, and Chen J. Molecular Structure of the Human CFTR Ion Channel. Cell 169: 85-95 e88, 2017. PMID: 28340353.
- Lohi H, Kujala M, Kerkela E, Saarialho-Kere U, Kestila M, and Kere J. Mapping of five new putative anion transporter genes in human and characterization of SLC26A6, a candidate gene for pancreatic anion exchanger. Genomics 70: 102-112, 2000. PMID: 11087667.
- Loiselle FB, Morgan PE, Alvarez BV, and Casey JR. Regulation of the human NBC3 Na+/HCO3- cotransporter by carbonic anhydrase II and PKA. Am J Physiol Cell Physiol 286: C1423-1433, 2004. PMID: 14736710.
- Luo X, Zheng W, Yan M, Lee MG, and Muallem S. Multiple functional P2X and P2Y receptors in the luminal and basolateral membranes of pancreatic duct cells. Am J Physiol 277: C205-215, 1999. PMID: 10444396.
- Luscher BP, Vachel L, Ohana E, and Muallem S. Cl(-) as a bona fide signaling ion. Am J Physiol Cell Physiol 318: C125-C136, 2020. PMID: 31693396.
- Ma T, Song Y, Gillespie A, Carlson EJ, Epstein CJ, and Verkman AS. Defective secretion of saliva in transgenic mice lacking aquaporin-5 water channels. J Biol Chem 274: 20071-20074, 1999. PMID: 10400615.
- Madden ME, and Sarras MP, Jr. Distribution of Na+,K+-ATPase in rat exocrine pancreas as monitored by K+-NPPase cytochemistry and [3H]-ouabain binding: a plasma membrane protein found primarily to be ductal cell associated. J Histochem Cytochem 35: 1365-1374, 1987. PMID: 2824600.
- Marteau C, Blanc G, Devaux MA, Portugal H, and Gerolami A. Influence of pancreatic ducts on saturation of juice with calcium carbonate in dogs. Dig Dis Sci 38: 2090-2097, 1993. PMID: 8223086.
- Martinez JR. Ion transport and water movement. J Dent Res 66 Spec No: 638-647, 1987. PMID: 3305642.
- McKenna R, and Frost SC. Overview of the carbonic anhydrase family. Subcell Biochem 75: 3-5, 2014. PMID: 24146371.
- Melvin JE, Yule D, Shuttleworth T, and Begenisich T. Regulation of fluid and electrolyte secretion in salivary gland acinar cells. Annu Rev Physiol 67: 445-469, 2005. PMID: 15709965.
- Messenger SW, Falkowski MA, and Groblewski GE. Ca(2)(+)-regulated secretory granule exocytosis in pancreatic and parotid acinar cells. Cell Calcium 55: 369-375, 2014. PMID: 24742357.
- Min X, Lee BH, Cobb MH, and Goldsmith EJ. Crystal structure of the kinase domain of WNK1, a kinase that causes a hereditary form of hypertension. Structure 12: 1303-1311, 2004. PMID: 15242606.
- Muhammad E, Leventhal N, Parvari G, Hanukoglu A, Hanukoglu I, Chalifa-Caspi V, Feinstein Y, Weinbrand J, Jacoby H, Manor E, Nagar T, Beck JC, Sheffield VC, Hershkovitz E, and Parvari R. Autosomal recessive hyponatremia due to isolated salt wasting in sweat associated with a mutation in the active site of Carbonic Anhydrase 12. Hum Genet 129: 397-405, 2011. PMID: 21184099.
- Namkung W, Han W, Luo X, Muallem S, Cho KH, Kim KH, and Lee MG. Protease-activated receptor 2 exerts local protection and mediates some systemic complications in acute pancreatitis. Gastroenterology 126: 1844-1859, 2004. PMID: 15188179.
- Namkung W, Lee JA, Ahn W, Han W, Kwon SW, Ahn DS, Kim KH, and Lee MG. Ca2+ activates cystic fibrosis transmembrane conductance regulator- and Cl- -dependent HCO3 transport in pancreatic duct cells. J Biol Chem 278: 200-207, 2003. PMID: 12409301.
- Namkung W, Yoon JS, Kim KH, and Lee MG. PAR2 exerts local protection against acute pancreatitis via modulation of MAP kinase and MAP kinase phosphatase signaling. Am J Physiol Gastrointest Liver Physiol 295: G886-894, 2008. PMID: 18755806.
- Nguyen TD, Meichle S, Kim US, Wong T, and Moody MW. P2Y(11), a purinergic receptor acting via cAMP, mediates secretion by pancreatic duct epithelial cells. Am J Physiol Gastrointest Liver Physiol 280: G795-804, 2001. PMID: 11292586.
- Nguyen TD, Moody MW, Steinhoff M, Okolo C, Koh DS, and Bunnett NW. Trypsin activates pancreatic duct epithelial cell ion channels through proteinase-activated receptor-2. J Clin Invest 103: 261-269, 1999. PMID: 9916138.
- North RA. Molecular physiology of P2X receptors. Physiol Rev 82: 1013-1067, 2002. PMID: 12270951.
- Novak I. ATP as a signaling molecule: the exocrine focus. News Physiol Sci 18: 12-17, 2003. PMID: 12531926.
- Novak I. Purinergic receptors in the endocrine and exocrine pancreas. Purinergic Signal 4: 237-253, 2008. PMID: 18368520.
- Novak I, and Greger R. Electrophysiological study of transport systems in isolated perfused pancreatic ducts: properties of the basolateral membrane. Pflugers Arch 411: 58-68, 1988. PMID: 3353213.
- Ohana E, Yang D, Shcheynikov N, and Muallem S. Diverse transport modes by the solute carrier 26 family of anion transporters. J Physiol 587: 2179-2185, 2009. PMID: 19015189.
- Ousingsawat J, Martins JR, Schreiber R, Rock JR, Harfe BD, and Kunzelmann K. Loss of TMEM16A causes a defect in epithelial Ca2+-dependent chloride transport. J Biol Chem 284: 28698-28703, 2009. PMID: 19679661.
- Pallagi P, Hegyi P, and Rakonczay Z, Jr. The Physiology and Pathophysiology of Pancreatic Ductal Secretion: The Background for Clinicians. Pancreas 44: 1211-1233, 2015. PMID: 26465950.
- Park HW, Nam JH, Kim JY, Namkung W, Yoon JS, Lee JS, Kim KS, Venglovecz V, Gray MA, Kim KH, and Lee MG. Dynamic regulation of CFTR bicarbonate permeability by [Cl-]i and its role in pancreatic bicarbonate secretion. Gastroenterology 139: 620-631, 2010. PMID: 20398666.
- Park M, Ko SB, Choi JY, Muallem G, Thomas PJ, Pushkin A, Lee MS, Kim JY, Lee MG, Muallem S, and Kurtz I. The cystic fibrosis transmembrane conductance regulator interacts with and regulates the activity of the HCO3- salvage transporter human Na+-HCO3- cotransport isoform 3. J Biol Chem 277: 50503-50509, 2002. PMID: 12403779.
- Park S, Hong JH, Ohana E, and Muallem S. The WNK/SPAK and IRBIT/PP1 pathways in epithelial fluid and electrolyte transport. Physiology (Bethesda) 27: 291-299, 2012. PMID: 23026752.
- Park S, Shcheynikov N, Hong JH, Zheng C, Suh SH, Kawaai K, Ando H, Mizutani A, Abe T, Kiyonari H, Seki G, Yule D, Mikoshiba K, and Muallem S. Irbit mediates synergy between ca(2+) and cAMP signaling pathways during epithelial transport in mice. Gastroenterology 145: 232-241, 2013. PMID: 23542070.
- Petersen OH. Stimulus-excitation coupling in plasma membranes of pancreatic acinar cells. Biochim Biophys Acta 694: 163-184, 1982. PMID: 6128029.
- Petersen OH, and Tepikin AV. Polarized calcium signaling in exocrine gland cells. Annu Rev Physiol 70: 273-299, 2008. PMID: 17850212.
- Piala AT, Moon TM, Akella R, He H, Cobb MH, and Goldsmith EJ. Chloride sensing by WNK1 involves inhibition of autophosphorylation. Sci Signal 7: ra41, 2014. PMID: 24803536.
- Poulsen JH, Fischer H, Illek B, and Machen TE. Bicarbonate conductance and pH regulatory capability of cystic fibrosis transmembrane conductance regulator. Proc Natl Acad Sci U S A 91: 5340-5344, 1994. PMID: 7515498.
- Preshaw RM, Cooke AR, and Grossman MI. Quantitative aspects of response of canine pancreas to duodenal acidification. Am J Physiol 210: 629-634, 1966. PMID: 5933217.
- Pushkin A, Abuladze N, Lee I, Newman D, Hwang J, and Kurtz I. Cloning, tissue distribution, genomic organization, and functional characterization of NBC3, a new member of the sodium bicarbonate cotransporter family. J Biol Chem 274: 16569-16575, 1999. PMID: 10347222.
- Quinton PM. Cystic fibrosis: impaired bicarbonate secretion and mucoviscidosis. Lancet 372: 415-417, 2008. PMID: 18675692.
- Quinton PM. The neglected ion: HCO3. Nat Med 7: 292-293, 2001. PMID: 11231624.
- Riordan JR, Rommens JM, Kerem B, Alon N, Rozmahel R, Grzelczak Z, Zielenski J, Lok S, Plavsic N, Chou JL, and et al. Identification of the cystic fibrosis gene: cloning and characterization of complementary DNA. Science 245: 1066-1073, 1989. PMID: 2475911.
- Rommens JM, Iannuzzi MC, Kerem B, Drumm ML, Melmer G, Dean M, Rozmahel R, Cole JL, Kennedy D, Hidaka N, and et al. Identification of the cystic fibrosis gene: chromosome walking and jumping. Science 245: 1059-1065, 1989. PMID: 2772657.
- Roos A, and Boron WF. Intracellular pH. Physiol Rev 61: 296-434, 1981. PMID: 7012859.
- Roussa E. Channels and transporters in salivary glands. Cell Tissue Res 343: 263-287, 2011. PMID: 21120532.
- Schaffalitzky de Muckadell OB, Fahrenkrug J, Watt-Boolsen S, and Worning H. Pancreatic response and plasma secretin concentration during infusion of low dose secretin in man. Scand J Gastroenterol 13: 305-311, 1978. PMID: 755275.
- Schroeder BC, Cheng T, Jan YN, and Jan LY. Expression cloning of TMEM16A as a calcium-activated chloride channel subunit. Cell 134: 1019-1029, 2008. PMID: 18805094.
- Shcheynikov N, Kim KH, Kim KM, Dorwart MR, Ko SB, Goto H, Naruse S, Thomas PJ, and Muallem S. Dynamic control of cystic fibrosis transmembrane conductance regulator Cl(-)/HCO3(-) selectivity by external Cl(-). J Biol Chem 279: 21857-21865, 2004. PMID: 15010471.
- Shcheynikov N, Son A, Hong JH, Yamazaki O, Ohana E, Kurtz I, Shin DM, and Muallem S. Intracellular Cl- as a signaling ion that potently regulates Na+/HCO3- transporters. Proc Natl Acad Sci U S A 112: E329-337, 2015. PMID: 25561556.
- Shcheynikov N, Wang Y, Park M, Ko SB, Dorwart M, Naruse S, Thomas PJ, and Muallem S. Coupling modes and stoichiometry of Cl-/HCO3- exchange by slc26a3 and slc26a6. J Gen Physiol 127: 511-524, 2006. PMID: 16606687.
- Shcheynikov N, Yang D, Wang Y, Zeng W, Karniski LP, So I, Wall SM, and Muallem S. The Slc26a4 transporter functions as an electroneutral Cl-/I-/HCO3- exchanger: role of Slc26a4 and Slc26a6 in I- and HCO3- secretion and in regulation of CFTR in the parotid duct. J Physiol 586: 3813-3824, 2008. PMID: 18565999.
- Shekarabi M, Zhang J, Khanna AR, Ellison DH, Delpire E, and Kahle KT. WNK Kinase Signaling in Ion Homeostasis and Human Disease. Cell Metab 25: 285-299, 2017. PMID: 28178566.
- Shin DH, Kim M, Kim Y, Jun I, Jung J, Nam JH, Cheng MH, and Lee MG. Bicarbonate permeation through anion channels: its role in health and disease. Pflugers Arch 472: 1003-1018, 2020. PMID: 32621085.
- Shirakabe K, Priori G, Yamada H, Ando H, Horita S, Fujita T, Fujimoto I, Mizutani A, Seki G, and Mikoshiba K. IRBIT, an inositol 1,4,5-trisphosphate receptor-binding protein, specifically binds to and activates pancreas-type Na+/HCO3- cotransporter 1 (pNBC1). Proc Natl Acad Sci U S A 103: 9542-9547, 2006. PMID: 16769890.
- Short DB, Trotter KW, Reczek D, Kreda SM, Bretscher A, Boucher RC, Stutts MJ, and Milgram SL. An apical PDZ protein anchors the cystic fibrosis transmembrane conductance regulator to the cytoskeleton. J Biol Chem 273: 19797-19801, 1998. PMID: 9677412.
- Smith ZD, Caplan MJ, Forbush B, 3rd, and Jamieson JD. Monoclonal antibody localization of Na+-K+-ATPase in the exocrine pancreas and parotid of the dog. Am J Physiol 253: G99-109, 1987. PMID: 2441610.
- Sohma Y, Gray MA, Imai Y, and Argent BE. HCO3- transport in a mathematical model of the pancreatic ductal epithelium. J Membr Biol 176: 77-100, 2000. PMID: 10882430.
- Sohma Y, Gray MA, Imai Y, and Argent BE. A mathematical model of the pancreatic ductal epithelium. J Membr Biol 154: 53-67, 1996. PMID: 8881027.
- Steward MC, Ishiguro H, and Case RM. Mechanisms of bicarbonate secretion in the pancreatic duct. Annu Rev Physiol 67: 377-409, 2005. PMID: 15709963.
- Stewart AK, Yamamoto A, Nakakuki M, Kondo T, Alper SL, and Ishiguro H. Functional coupling of apical Cl-/HCO3- exchange with CFTR in stimulated HCO3- secretion by guinea pig interlobular pancreatic duct. Am J Physiol Gastrointest Liver Physiol 296: G1307-1317, 2009. PMID: 19342507.
- Suzuki J, Umeda M, Sims PJ, and Nagata S. Calcium-dependent phospholipid scrambling by TMEM16F. Nature 468: 834-838, 2010. PMID: 21107324.
- Terker AS, Zhang C, Erspamer KJ, Gamba G, Yang CL, and Ellison DH. Unique chloride-sensing properties of WNK4 permit the distal nephron to modulate potassium homeostasis. Kidney Int 89: 127-134, 2016. PMID: 26422504.
- Ulrich CD, 2nd, Holtmann M, and Miller LJ. Secretin and vasoactive intestinal peptide receptors: members of a unique family of G protein-coupled receptors. Gastroenterology 114: 382-397, 1998. PMID: 9453500.
- Vachel L, Shcheynikov N, Yamazaki O, Fremder M, Ohana E, Son A, Shin DM, Yamazaki-Nakazawa A, Yang CR, Knepper MA, and Muallem S. Modulation of Cl(-) signaling and ion transport by recruitment of kinases and phosphatases mediated by the regulatory protein IRBIT. Sci Signal 11: 2018. PMID: 30377224.
- Veel T, Villanger O, Holthe MR, Cragoe EJ, Jr., and Raeder MG. Na(+)-H+ exchange is not important for pancreatic HCO3- secretion in the pig. Acta Physiol Scand 144: 239-246, 1992. PMID: 1316712.
- Venglovecz V, Hegyi P, Rakonczay Z, Jr., Tiszlavicz L, Nardi A, Grunnet M, and Gray MA. Pathophysiological relevance of apical large-conductance Ca(2)+-activated potassium channels in pancreatic duct epithelial cells. Gut 60: 361-369, 2011. PMID: 20940280.
- Venglovecz V, Pallagi P, Kemeny LV, Balazs A, Balla Z, Becskehazi E, Gal E, Toth E, Zvara A, Puskas LG, Borka K, Sendler M, Lerch MM, Mayerle J, Kuhn JP, Rakonczay Z, Jr., and Hegyi P. The Importance of Aquaporin 1 in Pancreatitis and Its Relation to the CFTR Cl(-) Channel. Front Physiol 9: 854, 2018. PMID: 30050452.
- Venglovecz V, Rakonczay Z, Jr., Ozsvari B, Takacs T, Lonovics J, Varro A, Gray MA, Argent BE, and Hegyi P. Effects of bile acids on pancreatic ductal bicarbonate secretion in guinea pig. Gut 57: 1102-1112, 2008. PMID: 18303091.
- Vilas G, Krishnan D, Loganathan SK, Malhotra D, Liu L, Beggs MR, Gena P, Calamita G, Jung M, Zimmermann R, Tamma G, Casey JR, and Alexander RT. Increased water flux induced by an aquaporin-1/carbonic anhydrase II interaction. Mol Biol Cell 26: 1106-1118, 2015. PMDI: 25609088.
- Wang Y, Soyombo AA, Shcheynikov N, Zeng W, Dorwart M, Marino CR, Thomas PJ, and Muallem S. Slc26a6 regulates CFTR activity in vivo to determine pancreatic duct HCO3- secretion: relevance to cystic fibrosis. EMBO J 25: 5049-5057, 2006. PMID: 17053783.
- Willoughby D, and Cooper DM. Organization and Ca2+ regulation of adenylyl cyclases in cAMP microdomains. Physiol Rev 87: 965-1010, 2007. PMID: 17615394.
- Wilson FH, Disse-Nicodeme S, Choate KA, Ishikawa K, Nelson-Williams C, Desitter I, Gunel M, Milford DV, Lipkin GW, Achard JM, Feely MP, Dussol B, Berland Y, Unwin RJ, Mayan H, Simon DB, Farfel Z, Jeunemaitre X, and Lifton RP. Human hypertension caused by mutations in WNK kinases. Science 293: 1107-1112, 2001. PMDI: 11498583.
- Wizemann V, and Schulz I. Influence of amphotericin, amiloride, ionophores, and 2,4-dinitrophenol on the secretion of the isolated cat's pancreas. Pflugers Arch 339: 317-338, 1973. PMID: 4735613.
- Yang D, Li Q, So I, Huang CL, Ando H, Mizutani A, Seki G, Mikoshiba K, Thomas PJ, and Muallem S. IRBIT governs epithelial secretion in mice by antagonizing the WNK/SPAK kinase pathway. J Clin Invest 121: 956-965, 2011. PMID: 21317537.
- Yang D, Shcheynikov N, and Muallem S. IRBIT: it is everywhere. Neurochem Res 36: 1166-1174, 2011. PMID: 21152975.
- Yang D, Shcheynikov N, Zeng W, Ohana E, So I, Ando H, Mizutani A, Mikoshiba K, and Muallem S. IRBIT coordinates epithelial fluid and HCO3- secretion by stimulating the transporters pNBC1 and CFTR in the murine pancreatic duct. J Clin Invest 119: 193-202, 2009. PMID: 19033647.
- Yang YD, Cho H, Koo JY, Tak MH, Cho Y, Shim WS, Park SP, Lee J, Lee B, Kim BM, Raouf R, Shin YK, and Oh U. TMEM16A confers receptor-activated calcium-dependent chloride conductance. Nature 455: 1210-1215, 2008. PMID: 18724360.
- You CH, Rominger JM, and Chey WY. Potentiation effect of cholecystokinin-octapeptide on pancreatic bicarbonate secretion stimulated by a physiologic dose of secretin in humans. Gastroenterology 85: 40-45, 1983. PMID: 6303892.
- Zeng M, Szymczak M, Ahuja M, Zheng C, Yin H, Swaim W, Chiorini JA, Bridges RJ, and Muallem S. Restoration of CFTR Activity in Ducts Rescues Acinar Cell Function and Reduces Inflammation in Pancreatic and Salivary Glands of Mice. Gastroenterology 153: 1148-1159, 2017. PMID: 28634110.
- Zeng W, Lee MG, and Muallem S. Membrane-specific regulation of Cl- channels by purinergic receptors in rat submandibular gland acinar and duct cells. J Biol Chem 272: 32956-32965, 1997. PMID: 9407075.
- Zeng W, Lee MG, Yan M, Diaz J, Benjamin I, Marino CR, Kopito R, Freedman S, Cotton C, Muallem S, and Thomas P. Immuno and functional characterization of CFTR in submandibular and pancreatic acinar and duct cells. Am J Physiol 273: C442-455, 1997. PMID: 9277342.
- Zhao H, and Muallem S. Na+, K+, and Cl- transport in resting pancreatic acinar cells. J Gen Physiol 106: 1225-1242, 1995. PMID: 8786358.
- Zhao H, Star RA, and Muallem S. Membrane localization of H+ and HCO3- transporters in the rat pancreatic duct. J Gen Physiol 104: 57-85, 1994. PMID: 7964596.