
Entry Version:
Citation:
Pancreapedia: Exocrine Pancreas Knowledge Base, DOI: 10.3998/panc.2017.02
| Attachment | Size |
|---|---|
| 579.73 KB |
1. Introduction
It is now established that almost all vertebrate organs show dynamic replacement or turnover at the level of individual proteins, organelles and cells. This allows replacement of damaged components and also regulation of functional mass (138). Regulatory loops where the expression of a mitogen is modulated by a feedback mechanism are common to many tissues. This is consistent with the observation that many tissues show both autonomous and regulated growth (109). Furthermore, in vertebrates, cell division and cell size are independent processes.
Regulation of pancreatic size is similar to other organs and occurs through genetic programing and environmental influences (especially food intake) with both directed at assuring that the organ can carry out its function in an energy efficient manner. In most vertebrates, the Islets of Langerhans are scattered through the gland but make up only 2-3% of the volume. Acinar cells make up about 85% of volume and ducts 5%, so overall pancreatic size reflects the exocrine component. However, acinar cells make up a smaller fraction of total cells and nuclei (approximately 50%) as other cell types such as myofibroblasts are much smaller. In the adult, islet mass and exocrine mass appear to be regulated independently. Assessment of islet volume generally requires histological analysis and will not be further considered here.
2. Measurement of Pancreatic Size
Overall pancreatic size is expressed as cm3 if volume is measured or g if weighed. Because the pancreas increases in size with age, the weight or volume is usually normalized to body weight. Until recently the most common technique was dissecting and weighing the pancreas. This yields values for young adults, usually males, fed a control diet, of 8-12 mg pancreatic weight/g body weight for mice (22, 64, 120, 125, 143), 22.5 mg PW/gBW for Syrian Hamster (124), 2.5-4 mg PW/g BW for rats (25, 34, 40, 57, 123, 158), and 0.5-1.1 g PW/kg BW for adult humans (13, 127). Note that the relative pancreas size gets smaller as the animal gets larger. Smaller animals have a larger body surface area and higher metabolic rate and food consumption relative to body size. We can speculate that the higher food consumption requires more digestive enzymes and hence a larger pancreas.
Recently, imaging studies using CT or MRI have been used and give similar values for pancreatic size in humans to earlier autopsy studies (72, 99, 119, 126). A study in pigs validated the method by comparing the pancreatic volume obtained by MRI to water displacement after pancreas removal (141). A recent study adopted MRI to determine the volume of mouse pancreas which was similar to water displacement (108). The main advantage of the imaging techniques is that they are non-invasive and can be carried out repetitively. Also, the volume for pancreatic parenchyma and fat can be determined separately. Such studies have shown that both total pancreatic mass and pancreatic fat increase with increasing BMI in humans (119) and decrease in both type 1 and type 2 diabetes (37, 74, 83, 152). The technique has been adapted to mice by use of a micro-MRI with a 7 Tesla magnet and worked best with the RARE (Rapid Acquisition with Relaxation) protocol (108). In this study, mice were perfusion-fixed initially so further work is necessary to establish the technique in living mice. In another study the pancreas architecture including the islet component was carried out by micro magnetic resonance microscopy (44).
3. Prenatal Pancreatic Growth
We can divide the lifespan of the pancreas into the prenatal, postnatal to adolescence and adult phases. The prenatal phase is dominated by organogenesis in which outgrowth from the foregut produces undifferentiated branching tubules expressing the transcription factor Pdx1 (Pancreatic and duodenal homeobox 1). These cells particularly the buds at the end of the tubules are considered to be transient progenitor cells. They will develop into acini, islets and mature ducts under the influence of mesenchyme and a number of other transcriptional regulators (61). However, these progenitor cells are not totipotent and probably should not be considered stem cells. During development, cell division and differentiation results in an increase in organ size and at birth, or within a few days, the pancreatic cells are able to assume their physiological function in response to environmental stimuli which activate neurohormonal control mechanisms. Some aspects of the pancreas change in mammals with weaning when a shift from a high fat to a high carbohydrate or high protein diet occurs and these changes can be considered either completion of development or an environmental (dietary) adaptation. The size of the pancreas at birth depends on the original number of progenitor cells as ablation of part of these cells during embryogenesis by targeting diphtheria toxin to the Pdx1 expressing cells reduces the size of the pancreas at birth and through postnatal life (139). This implies that each progenitor cell can give rise to a fixed amount of pancreas. At the time of birth the rat pancreas weighs about 5-10 mg and the relative size is 3 -3.8 mg/g body weight (57, 134).
4. Postnatal pancreatic growth
In the period from birth to adulthood the pancreas grows roughly in proportion to body growth. This has been quantitated both by dissecting and weighing the pancreas - and by imaging using CT or MRI followed by volume calculation. In a detailed study in Wistar rats, Iovanna et al evaluated pancreatic weight, protein and RNA from birth to 90 days (57). Pancreas weight increased faster than body mass until reaching a peak at 35 days and then declined (Figure 1A). A similar pattern of pancreas weight increase can be derived from the data of Snook carried out using CD strain rats which showed a peak PW/BW at 28 days (133, 134). Using micro-MRI, Paredes et al (2014) showed a linear relation in mice between pancreas volume and body weight between 20 days and 9 months of age (Figure 1B). By contrast, Stanger et al demonstrated a hyperbolic relationship with pancreatic growth faster than body growth in the first three weeks of life with gradual slowing thereafter (139). These differences are not major and may reflect differences in species, strain, or diet. In mice postnatal growth is not dependent on two of the major pancreas regulators, CCK and secretin, as the pancreas in young adult mice is similar when these peptides or their receptors undergo gene deletion (64, 120, 125). Similarly, in rats, pancreatic growth with age is not altered by a CCK antagonist (161). By contrast the growth of the neonatal pancreas can be increased by glucocorticoids and thyroxine which probably reflects a role of these hormones on development (90). The human pancreas also shows a linear relationship between size and age from birth to 25 years after which there is little change in size in adults until it begins declining around age 60 (119). In humans, due to the larger size, imaging can distinguish fat from parenchymal tissue and from midlife on the amount of fat in the pancreas increases with a concomitant decrease in the mass of exocrine tissue. During the growth phase, the increase in pancreatic mass primarily reflects cell division and the size of individual cells changes little after the first few days of life (27).

Figure 1. Pancreatic and body weight as a function of age. (A). Data for rats from 1 day post natal to 87 days. (Replotted from (57)). (B). Data for mice showing pancreatic volume and body weight from 20 to 280 days (108).
During the postnatal growth phase, pancreatic DNA increases in parallel to pancreatic weight. Acinar cell size shows a transient increase around birth due to the cells being packed with zymogen granules before secretion matures but is otherwise relatively constant. In support of the concept that pancreatic growth is driven by cell proliferation a number of studies have evaluated 3H-thymidine incorporation by autoradiography or the incorporation of the thymidine analog BrdU visualized by immunohistochemistry. Other studies evaluated specific proteins expressed by cells undergoing mitogenesis including proliferating cell nuclear antigen (PCNA) and Ki67. Finally, some early studies counted cells undergoing mitosis to obtain the mitotic index. These approaches generally parallel each other although showing different absolute numbers where the percentage of cells expressing PCNA is higher than those synthesizing DNA. Almost all of the earlier studies up to the year 2000 or latter were carried out in rats. All reported measures showed a high rate of acinar cell division in the last week of gestation with the rate falling around birth and then increasing postnatally to a peak index of 5-20% from 5 to 20 days and then a decline starting around weaning to adult rates at 30-60 days (26, 27, 95, 131, 151). In adult rats and mice on a nutritionally sound diet the indices of proliferation are under 1% (26, 27, 89, 131). There is little detailed evidence on clonality but both acinar and islet cells arise from division of differentiated cells. One additional complication is that starting around day 20 in rats the number of binucleate cells increases to near 50% at 50 days and these cells synthesize little more DNA (103). In a study in mice polyploidy first began at 20 days and increased to 9% at 90 days (147). Recently, most studies of pancreatic growth and regeneration are being carried out in mice and immunostaining with antibody to Ki67 has become the most common way to show which cell types are proliferating. However, there is much less quantitative data over the life span in mice, compared to rats.
5. Adaptive Pancreatic Growth
Need for regulation of pancreatic size
As discussed earlier, the size of the pancreas and the spectrum of digestive enzymes that it produces are generally related to the animal’s size and diet. The pancreas must provide sufficient enzymes for digestion and for trypsin a specific feedback system mediated by CCK is present to maintain the activity of trypsin in the intestinal lumen (69). Thus, the conditions that induce adaptive pancreatic growth are generally related to changes in diet. An increase in food intake such as occurs in pregnancy, lactation and cold exposure leads to an increase in pancreatic size in rodents (53, 61, 86, 92). Whether heavy exercise, which can double caloric intake, would affect pancreatic size would also be worthy of study. Also, there is little information as to whether pancreatic size adjusts to increased caloric consumption in humans.
The converse of increased food intake is starvation. Starvation is accompanied by loss of body and organ weights in all species examined from fish to mammals (85). While the pancreas has not always been evaluated, all organs in the GI tract decrease their weight during starvation especially the liver and intestine. In rats starved for 4 or 7 days, pancreatic weight decreased slightly more than body weight (84, 97, 148). Tissue weight and protein decreased more than DNA (93) consistent with cellular atrophy. Smaller acini with fewer ZG have also been observed by electron microscopy with starvation (62, 100, 150). This was most prominent in phase 3 of starvation when fat reserves have been utilized. Similar results have been reported in catfish (160) and sparrows (16). A study in obese humans who were fasted for 20 days showed a decrease in pancreatic juice volume and enzyme content upon pancreatic function testing consistent with pancreatic atrophy although pancreatic size was not measured (33). An extreme case is seen in snakes such as pythons that feed on a large meal and then carry out an extended fast (129). In this case, the involution of the GI tract occurs while the animal has adequate nutrients but food is not entering the gut. That the latter is important is shown by the results of parenteral feeding in rats where the intestine and pancreas atrophy even though normal nutrition is maintained (5, 30). Total pancreatic weight and protein were decreased in TPN while results for DNA were not clear. This effect of total parental nutrition (TPN) has been ascribed to the loss of GI hormones that normally would be released in response to feeding as well as the absence of nutrients in the lumen. Both CCK and bombesin have been shown to reduce the pancreatic atrophy induced by TPN (29, 111) and the atrophy was shown to be due to the loss of luminal protein as the combination of luminal fat, carbohydrate and amino acids could not reverse the pancreatic atrophy induced by TPN (4).
Another case of altered pancreatic function as a result of altered gut function is celiac disease. In celiac disease, ingestion of gluten, present in wheat and some other grains, triggers an immune attack on the duodenal mucosa which loses its villi resulting in a decreased surface area leading to malabsorption. Twenty to thirty percent of patients showed reduced exocrine pancreatic function and a portion of these show pancreatic insufficiency (28, 35, 115). While the mechanism is not clear, the most favored explanation is a reduction of GI hormone secretion, especially CCK (80). Other postulated mechanisms include reduced response to CCK, amino acid deficiency, and protein malnutrition. There is no evidence at present, however, that the pancreas is smaller in celiac disease.
Models for studying adaptive growth
Exogenous secretagogue stimulation
A large number of studies have shown that exogenous CCK can stimulate pancreatic growth. Because of the short half-life of CCK in vivo, CCK or its analogue caerulein have been given as 2 or 3 subcutaneous injections per day or by continuous infusion. Studies in rats mice and hamsters all show an increase in pancreatic size of 30-50% (1, 34, 101, 110, 124, 136). Secretin administration potentiates the increase in response to CCK in some but not all studies (51, 97, 98, 136). Somatostatin inhibits growth stimulated by CCK (123). This secretagogue driven increase in pancreatic mass was accompanied by increased thymidine incorporation into acinar cells peaking at 2-3 days accompanied by an increase in pancreatic DNA content (137). However, higher doses of CCK can induce apoptosis and a reduction in pancreatic weight (107, 144). The trophic action of CCK is mediated through CCK1 (CCK-A) receptors (113).
The effect of exogenous administration of other secretagogues has also been studied. Bombesin administration to rat, mouse and rabbit induced an increase in pancreatic wet weight and protein with or without an increase in DNA (23, 60, 67, 128). Because bombesin can stimulate the release of CCK several studies showed that the growth effect of bombesin was not blocked by a CCK antagonist which blocked the effect of exogenous CCK (10, 128). The effects of gastrin on pancreatic growth has been studied, but no consistent effect has emerged (90, 102, 135). Secretin by itself has no effect but may potentiate the action of CCK. The effect of administration of cholinergic analogs such as bethanechol and carbachol have been studied and shown to induce modest pancreatic growth which is less than the response to CCK (14, 92). However, it is not clear whether this effect is direct on acinar cells or mediated by release of some other regulator.
Although having considerable differences from in vivo pancreatic growth, primary cultures of acinar and duct cells have shown that CCK can directly stimulate acinar cell growth. Caerulein, CCK, and gastrin stimulated 3H-thymidine incorporation into monolayer cultures of mouse acinar cells on a collagen matrix (Figure 2) while carbachol, bombesin and substance P had no effect (77, 79). The effect involved both high and low affinity CCK receptors and could be inhibited by TGF-β (56, 78). CCK has also been shown to stimulate the growth of pancreatic cancer cell lines (132). Growth stimulatory effects of IGF-1 on mouse cells cultured on a laminin coated membrane have also been reported (145). However, the acinar cells in these cultures appear to have dedifferentiated (47) so they may be more of a model for recovery from damage than adaptive growth. Similar cultures of isolated duct cells have been studied but only growth factors and not secretagogues stimulated their growth (6).

Figure 2. Stimulation of mitogenesis in pancreatic acinar cells in monolayer culture on a collagen substrate. Top panels show thymidine incorporation into DNA when cells are stimulated by different secretagogues and growth factors. Bottom panels (C-E) show BrdU incorporation visualized by immunofluorescence. From (47).
Trypsin inhibitor feeding
It has been known for many years that chicks and rats fed raw soybean flour developed large pancreases with increased capacity to secrete digestive enzymes (9, 17, 82, 104). This was found due to the presence of active trypsin inhibitor (TI) and the effect could be reproduced by a variety of purified TIs (87, 116) and in a variety of animal species, but with most subsequent work in rats (54, 104, 159). Green and Lyman (42) showed that soybean trypsin inhibitor (SBTI) stimulated pancreatic secretion and presented evidence for a feedback mechanism in which active trypsin in the gut lumen inhibited pancreatic secretion. Thus, the feedback loop works to maintain an adequate level of trypsin and other proteases in the intestinal lumen. With the development of sensitive and specific CCK assays it was shown that release of CCK is part of the feedback loop (39, 70). Diversion of bile pancreatic juice also stimulates CCK release and pancreatic growth by removing active trypsin from the duodenum (2, 18, 38, 117, 146). Further information and references on this feedback loop can be found in (69, 76).
The feeding of either natural or orally active chemical trypsin inhibitors such as camostat (also known as FOY-305) can be used as a model to stimulate CCK driven pancreatic adaptive growth. The effects are bigger than the response to exogenous CCK which will stimulate pancreatic growth when given 2 or 3 times a day (24, 101, 136) probably because of desensitization to large exogenous boluses of CCK. Rodent and hamster pancreases will double in size in about a week in response to feeding TI (Figure 3) and the growth consists of hypertrophy and hyperplasia with the relative amount dependent on species. Current evidence suggests islet morphology and insulin content of the pancreas are unaffected (94). Co-administration of CCK antagonists block the effect of TI (25, 155). In rats the action of CCK was shown to be directly on acinar cells (158). In mice the effect of TI is not seen following the genetic deletion of either CCK or the CCK receptor (64, 125, 143). In both rats and mice the adaptive growth is reversible and when the TI is removed the pancreas returns to its original size (21, 105, 106). No dependence on secretin was seen in mice as TI had similar effects in mice where secretin signaling was genetically deleted (120). The evidence as to whether such a feedback loop involving CCK occurs in humans is mixed and may involve proteases other than trypsin (36, 72, 73).

Figure 3. Adaptive growth of mouse pancreas and change in plasma CCK in response to feeding a chemical trypsin inhibitor, camostat. Data from (143).
High protein diet
Consumption of a high protein diet is well established to cause pancreatic growth although most of the information is from rodents. Switching from a 5-10% casein diet to one containing 70-75% casein caused an increase in pancreatic weight, protein and DNA content reaching a plateau in 7-14 days (40, 41, 91). This was accompanied by a transient increase in plasma CCK and the growth response could be partially blocked with a CCK antagonist (41, 91). Protein is a primary stimulus for CCK release in the rat with casein being the most potent protein tested after trypsin inhibitor (71). The above experiments led to the concept that protein stimulated pancreatic growth through CCK and that this growth involved both hypertrophy and hyperplasia of acinar cells and was accompanied by an increase in pancreatic protease content.
However, it has been demonstrated that feeding a diet high in amino acids to rats stimulated pancreatic growth without stimulating CCK release (52, 71). This growth was also not blocked by a CCK antagonist. More definitively, a CCK deficient mouse fed a high protein diet showed an increase in pancreatic weight and protein content and a smaller increase in DNA (Figure 4). That the growth in CCK deficient mice was primarily due to cellular hypertrophy was shown by the increase in protein to DNA ratio, an increased acinar cell size and no increase in DNA synthesis as assessed by BrdU incorporation (22).

Figure 4. Effect of a high protein diet on pancreatic size and composition in normal and CCK deficient mice. Changes are shown for pancreas weight (A), Protein (B), DNA (C) and protein to DNA ratio (D) after 7 days on normal or high protein diet. From (22).
The converse of feeding a high protein diet is feeding a protein free or low protein diet in the presence of adequate calories. Normal rodent chow contains 18-22% protein, most often casein but what should be the normal amount is not always clear. Rat studies of high protein feeding have most often used 5 or 10% protein as the control or low protein to maximize the effect of the high protein. In rats on 5% protein the body weight failed to increase over the 14 day experimental period and the pancreas weight decreased slightly (41). In rats fed trypsin inhibitor or injected with caerulein to stimulate pancreatic growth, there was no pancreatic growth in animals on the 5% protein diet (41, 43). It has also been shown that gavage feeding of 9% casein increased plasma CCK but that 2% did not (71). Thus, rodents on diets with less than 5% protein may be compromised due to shortage of amino acids and lack of secretion of the pancreatic trophic hormone CCK. Rats feed a protein free diet lost pancreatic weight in excess of body weight and there was an increased apoptosis of acinar cells (32). A more detailed study has been carried out in mice where feeding protein-free diet was restricted to 4 days at which time there was an 11% decrease in body weight, no change in heart weight, but pancreas weight decreased by 24% (19). Most strikingly the protein content of the pancreas decreased almost 60 % while there was little fall in DNA (Figure 5). Electron microscopy showed decreased acinar size and a large decrease in the number of zymogen granules in mice, rats, rabbits and monkeys (19, 66, 114, 149). The changes were reversible and interpreted as cellular atrophy. Small sized acinar cells and failure of digestion has also been reported for children with Kwashiorkor (7, 112, 153). This was interpreted as a failure of the pancreas to secrete digestive enzymes.
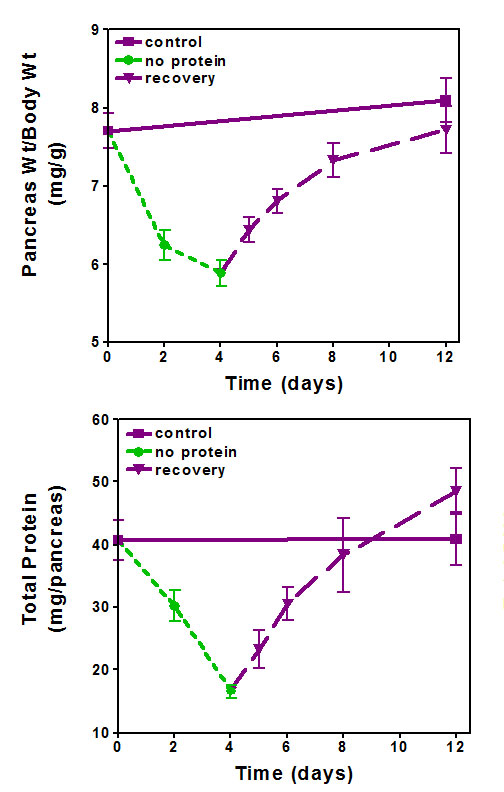
Figure 5. Feeding a protein free diet induces pancreatic atrophy as shown by a decrease in pancreatic weight (A) and protein content (B). From (19).
6. Signaling Pathways Mediating Adaptive growth
Pathways regulating cell size
The primary pathway regulating adaptive cell size growth is centered on the mechanistic target of rapamycin complex 1 (mTORC1) (31, 65, 156). This pathway was discovered in yeast where TOR1 and TOR2 were identified as the targets for the macrolide rapamycin which was isolated from a soil bacterium discovered on Rapa Nui (Easter Island) and found to inhibit growth. Mammals have one form of TOR named mTOR (m originally stood for mammalian but now for mechanistic) which forms two distinct complexes; mTORC1 being primarily related to growth and is sensitive to rapamycin while mTORC2 regulates the actin cytoskeleton and is relatively insensitive to rapamycin. Each complex has a number of additional regulatory proteins. TORC1 contains six known proteins with the scaffolding protein raptor, and the inhibitory protein, proline-rich Akt substrate of 40 kDa (Pras40) being unique to mTORC1. Other subunits are shared with mTORC2 which also contains its own unique protein, rictor. mTOR the catalytic subunit is a 280 kDa protein that belongs to the phosphoinositide 3-kinase related kinase family and contains multiple domains including one binding FKBP-12, the receptor for both FK506 and rapamycin. The mTORC1 pathway integrates input from growth factors, energy status, oxygen, and amino acids. When these are present mTORC1 stimulates protein synthesis, lipid synthesis, cell cycle progression and inhibits autophagy (65, 130).
A diagram of the pathway as known for pancreatic acinar cells is shown without all details in Fig 6. The CCK1 receptor, M3 muscarinic receptors and insulin and other growth factor receptors all activate phosphatidyl inositol 3-kinase (PI3-K). PI3-K phosphorylates and activates Akt also known as PKB which phosphorylates and inactivates the tuberous sclerosis complex (TSC). In other cells hypoxia and low cellular energy status are known to activate TSC through REDD and AMPK respectively. The TSC complex inhibits activation of the G protein Rheb which when liganded by GTP activates TORC1 (68). This activation process in cells that have been studied involves migration to the surface of lysosomes and is the site where amino acids, especially leucine and arginine activate TORC1 by a process involving a number of other proteins (3, 15). TORC1 in pancreatic acinar cells is known to be activated by branched chain amino acids (121).

Figure 6. Schematic drawing of mTORC1 signaling pathway in pancreatic acinar cells.
The best characterized substrates of TORC1 are ribosomal protein S6 kinase (S6K) 1 and initiation factor 4E binding protein 1 (4E-BP1). S6K1 phosphorylates a number of substrates including ribosomal small subunit 6 (S6) while phosphorylation of 4E-BP1 releases IF4E which binds the 5’ cap on mRNA to initiate cap dependent translation which accounts for about 90% of mRNA species. Both of these events are readily demonstrable in intact pancreas and pancreatic acini in response to CCK, growth factors and amino acids (11, 12, 20, 121). A third action in acini contributing to protein synthesis is the activation of elongation factor 2 (eEF2) (122).
The importance of TORC1 for adaptive pancreatic growth is shown by the fact that pancreatic growth stimulated by feeding TI is blocked by rapamycin (20) or by genetic deletion of raptor in acinar cells (S. Crozier and J.A. Williams, unpublished data) both of which selectively block the TORC1 pathway without affecting other pathways such as the activation of ERK and JNK leading to early response genes (20). Interestingly, rapamycin blocks adaptive growth but has little effect on the baseline size of the pancreas. By contrast, feeding a protein free diet blocks the TORC1 pathway and causes a reduction in pancreatic size due to cellular atrophy (19). This difference may be due to the fact that some actions of TORC1 are known to be insensitive to rapamycin (59). When raptor is deleted in acinar cells leading to a loss of TORC1 components, there is both an increase in apoptosis and a dedifferentiation of acinar cells. Thus, there is clearly more to learn of the importance of the TORC1 pathway in acinar cells. Although not presented in detail here, islet beta and alpha cells also are dependent on TORC1 to maintain their mass and demonstrate adaptive growth (157).
Pathways regulating pancreatic mitogenesis
Because of the prominent role of CCK in inducing adaptive pancreatic growth, attention has focused on how CCK receptors activate acinar cell mitogenesis. Mature acinar cells appear to exist in a Go state and mitogenic stimuli cause them to enter the cell cycle. Early response genes are a class of genes known to be rapidly upregulated following cellular stimulation and function as mediators coupling short-term signals to longer term cellular responses such as growth by regulating other genes (118). Initial work showed that CCK induced various early response genes in rat pancreas or isolated acini including c-fos, c-jun, Egr-1 and c-myc (58, 81). This led to enhanced activity of AP-1 transcription factor which in its most common form is a fos-jun heterodimer. A more complete study of the response of the mouse pancreas to feeding TI in chow evaluated mRNA for 18 early response genes with 17 showing a rapid increase with a maximum effect seen at 1 to 4 hours followed by a decline (Fig 7) (46). Protein expression for c-jun, c-fos, ATF-3, Egr-1 and JunB peaked at two hours. Immunohistochemistry showed nuclear localization and AP-1 activity was increased in nuclei. The effect of feeding TI was largely blocked by a CCK antagonist and there was no effect of refeeding regular chow. That this is a direct effect of CCK was supported by similar findings in monolayer cultures of acinar cells (47). In the in vitro study CCK also induced c-Jun, AP-1 activity and cyclin D1 expression. Dominant negative JNK or shRNA blocked the c-Jun increase and prevented enhanced mitogenesis. In this culture system acinar cells dedifferentiate before mitogenesis, so it is not a perfect model but shows that CCK can directly stimulate mitogenesis as shown earlier by Logsdon (77, 79).
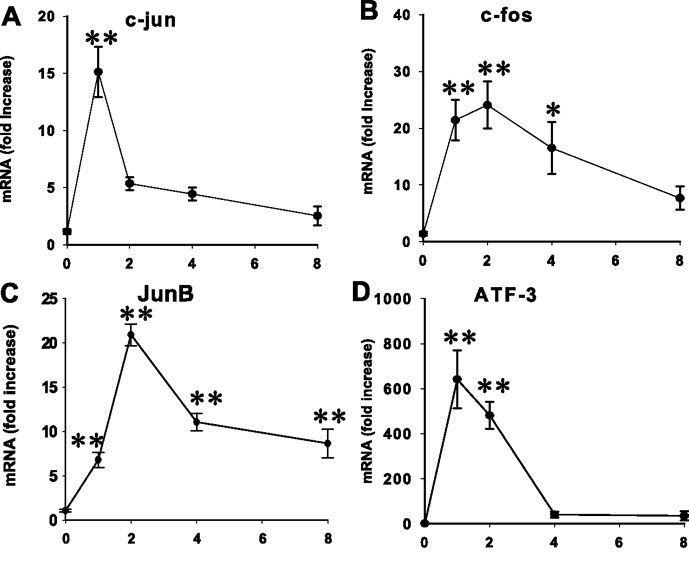
Figure 7. Expression of four early response genes peaking at 1 or 2 hours after refeeding chow containing trypsin inhibitor to mice after an overnight fast. From (46).
Calcineurin-NFAT pathway
CCK is known to activate a number of signaling pathways but a central role has been assigned to activation of intracellular Ca2+ signaling for the stimulation of digestive enzyme secretion. A sustained increase in Ca2+ is known to activate the Ca2+ regulated phosphatase, calcineurin (CN) in a variety of cells. Utilizing the pharmacological inhibitors cyclosporine A (CsA) and FK-506, Tashiro et al showed that active CN was necessary for TI stimulated pancreatic growth that occurs by activation of mitogenesis (143). A major target of CN are NFATs (Nuclear Factor of Activated T Cells). In the basal state NFATs are phosphorylated and remain in the cytoplasm. Upon stimulation which increases intracellular Ca2+, activated CN dephosphorylates NFAT which move into the nucleus and either alone or in partnership with AP-1 activates specific gene expression (Fig 8). In isolated pancreatic acini, CCK stimulated NFAT translocation into nuclei with the effect dependent on CN (49). An NFAT reporter was also used to show increased NFAT activity. Similar activation of NFATs with nuclear translocation was seen in vivo in response to feeding TI.
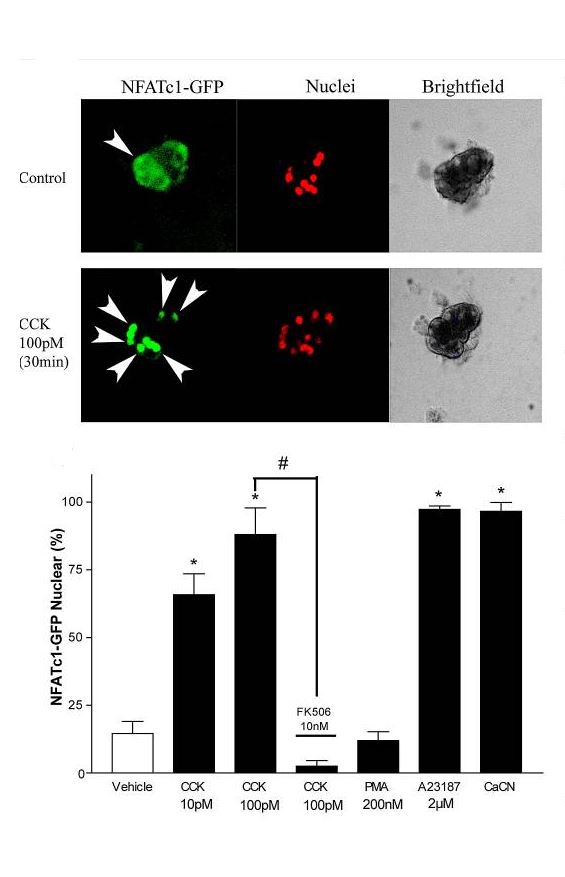
Figure 8. CCK stimulates nuclear translocation of NFATc1-GFP in mouse pancreatic acini. From (49).
Further evidence for the importance of CN came from the discovery that Rcan1 (Regulator of calcineurin 1) also known as DSCR1 was highly induced by feeding TI with the induction almost totally inhibitable by FK-506. Rcan1 blocked CN action in cell models and when induced in pancreatic acinar cells in vivo using a LSL Rcan1 construct activated by Ela-Cre completely blocked adaptive growth in response to feeding TI (Fig 9) (48). Thus, both pharmacological and genetic inhibition of CN blocked adaptive growth. Though it is reasonable to presume this is mediated by NFATs other CN substrates could also participate.
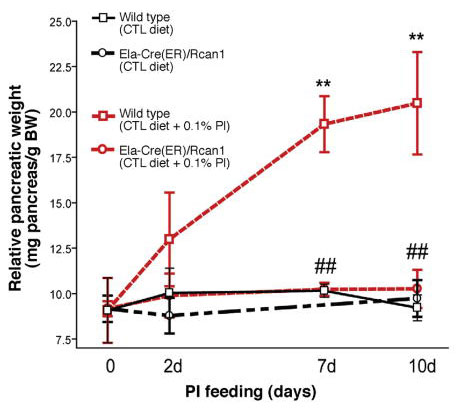
Figure 9. Overexpression of Rcan 1 in acinar cells blocks TI stimulated pancreatic growth. From (48).
RNA microarray analysis of pancreatic genes revealed 38 genes induced by TI two hours after feeding that were at least 70% inhibited by FK-506 (48). These changes were not observed in CCK deficient mice. The 38 genes included feedback inhibitors (Rcan1, Socs3, Rgs2, Socs 2, Lif), growth factors (FGF21, HBEGF) and transcriptional regulators. About 60% had predicted NFAT regulation and CHIP analysis confirmed this for 5 genes in pancreatic 266-6 acinar cells stimulated with calcium ionophore (48).
TORC1 Pathway
The TORC1 pathway discussed above as playing a role in the increased cell size seen in response to high protein feeding is also necessary for adaptive growth involving cell proliferation. This has been shown through the use of rapamycin which blocks both mitogenesis and the increase in pancreatic weight, protein and DNA in response to TI feeding (20). Genetically, deletion of raptor, an essential component of TORC1 also blocks adaptive growth (S. Crozier and J.A. Williams, unpublished data). Similarly mTORC1 is required for islet beta cell growth (8). In yeast, rapamycin blocks cell division because mitogenesis can not occur without the increase in cell protein required to convert one cell into two. This does not appear to be the case in mammalian cells and attention has focused instead on actions of TORC1 on cell cycle regulators such as Cyclin D1 and CDK4 which are sensitive to rapamycin.
MAPK Pathways
CCK and other pancreatic secretagogues activate multiple MAPK pathways including ERK, JNK and p38 MAPK (154). Of these both the ERK and JNK pathways are also activated by feeding TI (142). Interestingly the activation of JNK was blocked by FK-506 pretreatment while activation of ERK which occurred rapidly with peak activation at 30 min was unaffected. A latter study showed a more prolonged activation of ERK (55). JNK activation is known to be important for mitogenesis in cultured acinar cells (47) but has not been studied in normal animals. A requirement for ERK for adaptive growth in vivo in mice has been shown using new ERK inhibitors developed for in vivo administration as potential cancer therapeutic agents (Fig 10). PD-0325901 and GSK-11202012 (Trametenib) can be mixed in chow or gavage fed and inhibit by binding to the catalytic site of MEK thereby blocking ERK with low toxicity. The MEK inhibitors blocked pancreatic ERK signaling, adaptive growth, and the increased DNA synthesis stimulated by feeding TI to mice (55). Interestingly the MEK inhibitor blocked induction of c-Fos but had no effect on c-Jun induction. It also blocked the expression of cyclins D1, D3, E, and A along with PCNA (55). The ERK pathway has previously been shown to play a proliferative role in multiple other cells and tissues (88) although its mechanistic target is not clearly established.

Figure 10. Inhibition of ERK blocks pancreatic growth. Mice were fed TI for 5 days after gavaging with the ERK inhibitor PD0325901. From (55).
Stat3
Bioinformatics analysis of 318 genes which were upregulated in pancreas from 1 to 8 hours after feeding TI suggested involvement of Jak-STAT signaling (50). An increase in phospho STAT3 showed a strong nuclear occupancy peaking at 75% of acinar cell nuclei at 2 h. Western blotting showed a large increase in p-STAT3 without a change in total STAT3. Socs2 which acts as a negative feedback regulator of STAT3 is a transcriptional target of STAT3 and its mRNA was found to also be enhanced. Interestingly, in isolated acini CCK fails to activate Jak-STAT signaling while growth factors can. Since growth factors such as HB-EGF are induced by TI feeding, it is possible that CCK does not work directly but rather through a growth factor whose receptor can activate Jak-Stat signaling. At present, there is no direct evidence that STAT3 is involved in adaptive pancreatic growth. However, it seems likely that it will be involved as STAT3 is involved in the malignant transformation of acinar cells (45, 96). Also, STAT5 and Socs3 have been shown to regulate growth of pancreatic islet beta cells (75).
Overall model of mitogenesis
It seems clear that acinar cell mitogenesis is under different and more complex control than is secretion of digestive enzymes. Blockage of three different pathways, the calcineurin-NFAT, the TORC1 and the ERK pathway all block adaptive growth indicating that all must be activated for mitogenesis to occur (Figure 11). It is not clear whether all three converge on a single rate limiting step or that multiple key steps exist and each is activated by a different pathway. It appears that the induction of multiple cyclins is blocked by inhibiting each pathway so the regulated step is most likely connected with moving cells out of Go and initiating the cell cycle. In addition to completing the understanding of how acinar cell mitogenesis is initiated, work is needed on whether and how there is a parallel increase in ducts and blood vessels as the pancreas grows. Similar studies have also yielded an incomplete map for regulation of islet beta cell growth (63, 140).

Figure 11. Three CCK-activated signaling pathways required for pancreatic acinar cell growth by proliferation of acinar cells.
7. References
- Arnesjo, II and Lundquist I. Effects on exocrine and endocrine rat pancreas of long-term administration of CCK-PZ (cholecystokinin-pancreozymin) or synthetic octapeptide - CCK-PZ. Scand J Gastroenterol 11(5): 529-535,1976. PMID: 785590.
- Bamba T, Ishizuka Y and Hosoda S. Effect of intrinsic CCK and CCK antagonist on pancreatic growth and pancreatic enzyme secretion in pancreaticobiliary diversion rats. Dig Dis Sci 38(4): 653-659,1993. PMID: 7681747.
- Bar-Peled L and Sabatini DM. Regulation of mTORC1 by amino acids. Trends Cell Biol 24(7): 400-406,2014. PMID: 24698685.
- Baumler MD, Koopmann MC, Thomas DD, Ney DM and Groblewski GE. Intravenous or luminal amino acids are insufficient to maintain pancreatic growth and digestive enzyme expression in the absence of intact dietary protein. Am J Physiol Gastrointest Liver Physiol 299(2): G338-347,2010. PMID: 20539007.
- Baumler MD, Nelson DW, Ney DM and Groblewski GE. Loss of exocrine pancreatic stimulation during parenteral feeding suppresses digestive enzyme expression and induces Hsp70 expression. Am J Physiol Gastrointest Liver Physiol 292(3): G857-866,2007. PMID: 17138970.
- Bhattacharyya E, Panchal A, Wilkins TJ, de Ondarza J and Hootman SR. Insulin, transforming growth factors, and substrates modulate growth of guinea pig pancreatic duct cells in vitro. Gastroenterology 109(3): 944-952,1995. PMID: 7657124.
- Blackburn WR and Vinijchaikul K. The pancreas in kwashiorkor. An electron microscopic study. Lab Invest 20(4): 305-318,1969. PMID: 5776325.
- Blandino-Rosano M, Chen AY, Scheys JO, Alejandro EU, Gould AP, Taranukha T, et al. mTORC1 signaling and regulation of pancreatic beta-cell mass. Cell Cycle 11(10): 1892-1902,2012. PMID: 22544327.
- Booth AN, Robbins DJ, Ribelin WE, Deeds F, Smith AK and Rackis JJ. Prolonged Pancreatic Hypertrophy and Reversibility in Rats Fed Raw Soybean Meal. Proc Soc Exp Biol Med 116: 1067-1069,1964. PMID: 14230346.
- Borysewicz R, Ren KJ, Mokotoff M and Lee PC. Direct effect of bombesin on pancreatic and gastric growth in suckling rats. Regul Pept 41(2): 157-169,1992. PMID: 1438986.
- Bragado MJ, Groblewski GE and Williams JA. Regulation of protein synthesis by cholecystokinin in rat pancreatic acini involves PHAS-I and the p70 S6 kinase pathway. Gastroenterology 115(3): 733-742,1998. PMID: 9721171.
- Bragado MJ, Tashiro M and Williams JA. Regulation of the initiation of pancreatic digestive enzyme protein synthesis by cholecystokinin in rat pancreas in vivo. Gastroenterology 119(6): 1731-1739,2000. PMID: 11113094.
- Caglar V KB, Uygur R,Alkoc OA,Ozen OA and Demirel H. Study of Volume, Weight and Size of Normal Pancreas, Spleen and Kidney in Adults Autopsies. Forensic Medicine and Anatomy Research 2(3): 63-60,2014. PMID.
- Carling RC and Templeton D. The effect of carbachol and isoprenaline on cell division in the exocrine pancreas of the rat. Q J Exp Physiol 67(4): 577-585,1982. PMID: 6185966.
- Chantranupong L, Scaria SM, Saxton RA, Gygi MP, Shen K, Wyant GA, et al. The CASTOR Proteins Are Arginine Sensors for the mTORC1 Pathway. Cell 165(1): 153-164,2016. PMID: 26972053.
- Chediack JG, Funes SC, Cid FD, Filippa V and Caviedes-Vidal E. Effect of fasting on the structure and function of the gastrointestinal tract of house sparrows (Passer domesticus). Comp Biochem Physiol A Mol Integr Physiol 163(1): 103-110,2012. PMID: 22613787.
- Chernick SS, Lepkovsky S and Chaikoff IL. A dietary factor regulating the enzyme content of the pancreas; changes induced in size and proteolytic activity of the chick pancreas by the ingestion of raw soy-bean meal. Am J Physiol 155(1): 33-41,1948. PMID: 18102665.
- Chu M, Borch K, Lilja I, Blomqvist L, Rehfeld JF and Ihse I. Endogenous hypercholecystokininemia model in the hamster: trophic effect on the exocrine pancreas. Pancreas 7(2): 220-225,1992. PMID: 1553371.
- Crozier SJ, D'Alecy LG, Ernst SA, Ginsburg LE and Williams JA. Molecular mechanisms of pancreatic dysfunction induced by protein malnutrition. Gastroenterology 137(3): 1093-1101, 1101 e1091-1093,2009. PMID: 19427311.
- Crozier SJ, Sans MD, Guo L, D'Alecy LG and Williams JA. Activation of the mTOR signalling pathway is required for pancreatic growth in protease-inhibitor-fed mice. J Physiol 573(Pt 3): 775-786,2006. PMID: 16613881.
- Crozier SJ, Sans MD, Lang CH, D'Alecy LG, Ernst SA and Williams JA. CCK-induced pancreatic growth is not limited by mitogenic capacity in mice. Am J Physiol Gastrointest Liver Physiol 294(5): G1148-1157,2008. PMID: 18356533.
- Crozier SJ, Sans MD, Wang JY, Lentz SI, Ernst SA and Williams JA. CCK-independent mTORC1 activation during dietary protein-induced exocrine pancreas growth. Am J Physiol Gastrointest Liver Physiol 299(5): G1154-1163,2010. PMID: 20798356.
- Damge C, Hajri A, Lhoste E and Aprahamian M. Comparative effect of chronic bombesin, gastrin-releasing peptide and caerulein on the rat pancreas. Regul Pept 20(2): 141-150,1988. PMID: 2452459.
- Dembinski AB and Johnson LR. Stimulation of pancreatic growth by secretin, caerulein, and pentagastrin. Endocrinology 106(1): 323-328,1980. PMID: 7349962.
- Douglas BR, Woutersen RA, Jansen JB, Rovati LC and Lamers CB. Comparison of the effect of lorglumide on pancreatic growth stimulated by camostate in rat and hamster. Life Sci 46(4): 281-286,1990. PMID: 2304371.
- Elsässer H Adler G, and Kern HF. Replication and Regeneration of the Pancreas. The Pancreas: Biology, Pathobiology, and Disease. Ed by VLW Go et all, pp75-86, 1993.
- Elsasser HP, Biederbick A and Kern HF. Growth of rat pancreatic acinar cells quantitated with a monoclonal antibody against the proliferating cell nuclear antigen. Cell Tissue Res 276(3): 603-609,1994. PMID: 7914831.
- Evans KE, Leeds JS, Morley S and Sanders DS. Pancreatic insufficiency in adult celiac disease: do patients require long-term enzyme supplementation? Dig Dis Sci 55(10): 2999-3004,2010. PMID: 20458623.
- Fan BG, Axelson J, Sternby B, Rehfeld J, Ihse I and Ekelund M. Total parenteral nutrition affects the tropic effect of cholecystokinin on the exocrine pancreas. Scand J Gastroenterol 32(4): 380-386,1997. PMID: 9140162.
- Fan BG, Salehi A, Sternby B, Axelson J, Lundquist I, Andren-Sandberg A, et al. Total parenteral nutrition influences both endocrine and exocrine function of rat pancreas. Pancreas 15(2): 147-153,1997. PMID: 9260199.
- Fingar DC and Blenis J. Target of rapamycin (TOR): an integrator of nutrient and growth factor signals and coordinator of cell growth and cell cycle progression. Oncogene 23(18): 3151-3171,2004. PMID: 15094765.
- Fitzgerald PJ Herman LL, Carol B,Roque A,Marsh WH,Rosenstock L, et al. Pancreatic acinar cell regeneration. American Journal of Pathology 52(5): 983-1012,1968,1968. PMID: 2013346.
- Folsch UR, Dreessen UW, Talaulicar M, Willms B and Creutzfeldt W. Effect of long-term fasting of obese patients on pancreatic exocrine function, gastrointestinal hormones and bicarbonate concentration in plasma. Z Gastroenterol 22(7): 357-364,1984. PMID: 6207670.
- Folsch UR, Winckler K and Wormsley KG. Influence of repeated administration of cholecystokinin and secretin on the pancreas of the rat. Scand J Gastroenterol 13(6): 663-671,1978. PMID: 694411.
- Freeman HJ. Pancreatic endocrine and exocrine changes in celiac disease. World J Gastroenterol 13(47): 6344-6346,2007. PMID: 18081222.
- Friess H, Kleeff J, Isenmann R, Malfertheiner P and Buchler MW. Adaptation of the human pancreas to inhibition of luminal proteolytic activity. Gastroenterology 115(2): 388-396,1998. PMID: 9679044.
- Gaglia JL, Guimaraes AR, Harisinghani M, Turvey SE, Jackson R, Benoist C, et al. Noninvasive imaging of pancreatic islet inflammation in type 1A diabetes patients. J Clin Invest 121(1): 442-445,2011. PMID: 21123946.
- Gasslander T, Axelson J, Hakanson R, Ihse I, Lilja I and Rehfeld JF. Cholecystokinin is responsible for growth of the pancreas after pancreaticobiliary diversion in rats. Scand J Gastroenterol 25(10): 1060-1065,1990. PMID: 2263879.
- Goke B, Printz H, Koop I, Rausch U, Richter G, Arnold R, et al. Endogenous CCK release and pancreatic growth in rats after feeding a proteinase inhibitor (camostate). Pancreas 1(6): 509-515,1986. PMID: 3562444.
- Green GM, Jurkowska G, Berube FL, Rivard N, Guan D and Morisset J. Role of cholecystokinin in induction and maintenance of dietary protein-stimulated pancreatic growth. Am J Physiol 262(4 Pt 1): G740-746,1992. PMID: 1373568.
- Green GM, Levan VH and Liddle RA. Plasma cholecystokinin and pancreatic growth during adaptation to dietary protein. Am J Physiol 251(1 Pt 1): G70-74,1986. PMID: 3524262.
- Green GM and Lyman RL. Feedback regulation of pancreatic enzyme secretion as a mechanism for trypsin inhibitor-induced hypersecretion in rats. Proc Soc Exp Biol Med 140(1): 6-12,1972. PMID: 5033119.
- Green GM, Sarfati PD and Morisset J. Lack of effect of cerulein on pancreatic growth of rats fed a low-protein diet. Pancreas 6(2): 182-189,1991. PMID: 1715991.
- Grippo PJ, Venkatasubramanian PN, Knop RH, Heiferman DM, Iordanescu G, Melstrom LG, et al. Visualization of mouse pancreas architecture using MR microscopy. Am J Pathol 179(2): 610-618,2011. PMID: 21683673.
- Gruber R, Panayiotou R, Nye E, Spencer-Dene B, Stamp G and Behrens A. YAP1 and TAZ Control Pancreatic Cancer Initiation in Mice by Direct Up-regulation of JAK-STAT3 Signaling. Gastroenterology 151(3): 526-539,2016. PMID: 27215660.
- Guo L, Sans MD, Gurda GT, Lee SH, Ernst SA and Williams JA. Induction of early response genes in trypsin inhibitor-induced pancreatic growth. Am J Physiol Gastrointest Liver Physiol 292(2): G667-677,2007. PMID: 17095753.
- Guo L, Sans MD, Hou Y, Ernst SA and Williams JA. c-Jun/AP-1 is required for CCK-induced pancreatic acinar cell dedifferentiation and DNA synthesis in vitro. Am J Physiol Gastrointest Liver Physiol 302(12): G1381-1396,2012. PMID: 22461029.
- Gurda GT, Crozier SJ, Ji B, Ernst SA, Logsdon CD, Rothermel BA, et al. Regulator of calcineurin 1 controls growth plasticity of adult pancreas. Gastroenterology 139(2): 609-619, 619 e601-606,2010. PMID: 20438729.
- Gurda GT, Guo L, Lee SH, Molkentin JD and Williams JA. Cholecystokinin activates pancreatic calcineurin-NFAT signaling in vitro and in vivo. Mol Biol Cell 19(1): 198-206,2008. PMID: 17978097.
- Gurda GT, Wang JY, Guo L, Ernst SA and Williams JA. Profiling CCK-mediated pancreatic growth: the dynamic genetic program and the role of STATs as potential regulators. Physiol Genomics 44(1): 14-24,2012. PMID: 22010007.
- Haarstad H and Petersen H. The effects of graded doses of a cholecystokinin-like peptide with and without secretin on pancreatic growth and synthesis of RNA and polyamines in rats. Scand J Gastroenterol 24(8): 907-915,1989. PMID: 2480634.
- Hara H, Narakino H, Kiriyama S and Kasai T. Induction of pancreatic growth and proteases by feeding a high amino acid diet does not depend on cholecystokinin in rats. J Nutr 125(5): 1143-1149,1995. PMID: 7738673.
- Harada E and Kanno T. Progressive enhancement in the secretory functions of the digestive system of the rat in the course of cold acclimation. J Physiol 260(3): 629-645,1976. PMID: 978571.
- Hasdai A and Liener IE. Growth, digestibility and enzymatic activities in the pancreas and intestines of hamsters fed raw and heated soy flour. J Nutr 113(3): 662-668,1983. PMID: 6186799.
- Holtz BJ, Lodewyk KB, Sebolt-Leopold JS, Ernst SA and Williams JA. ERK activation is required for CCK-mediated pancreatic adaptive growth in mice. Am J Physiol Gastrointest Liver Physiol 307(7): G700-710,2014. PMID: 25104499.
- Hoshi H and Logsdon CD. Both low- and high-affinity CCK receptor states mediate trophic effects on rat pancreatic acinar cells. Am J Physiol 265(6 Pt 1): G1177-1181,1993. PMID: 8279569.
- Iovanna JL, Dusetti N, Cadenas B and Calvo EL. Changes in growth and pancreatic mRNA concentrations during postnatal development of rat pancreas. Pancreas 5(4): 421-426,1990. PMID: 1696380.
- Ji B, Chen XQ, Misek DE, Kuick R, Hanash S, Ernst S, et al. Pancreatic gene expression during the initiation of acute pancreatitis: identification of EGR-1 as a key regulator. Physiol Genomics 14(1): 59-72,2003. PMID: 12709512.
- Kang SA, Pacold ME, Cervantes CL, Lim D, Lou HJ, Ottina K, et al. mTORC1 phosphorylation sites encode their sensitivity to starvation and rapamycin. Science 341(6144): 1236566,2013. PMID: 23888043.
- Karkashan EM, MacNaughton WK and Gall DG. Evidence of a physiological role for bombesin in the postnatal development of the rabbit pancreas. Biol Neonate 62(6): 395-401,1992. PMID: 1282037.
- Kim YS, Kim WJ, Kim HK and Hong SS. Effect of cold and hot environments on the exocrine pancreas of rats. Yonsei Med J 11(1): 1-9,1970. PMID: 5516840.
- Kitagawa T and Ono K. Ultrastructure of pancreatic exocrine cells of the rat during starvation. Histol Histopathol 1(1): 49-57,1986. PMID: 2980101.
- Kulkarni RN, Mizrachi EB, Ocana AG and Stewart AF. Human beta-cell proliferation and intracellular signaling: driving in the dark without a road map. Diabetes 61(9): 2205-2213,2012. PMID: 22751699.
- Lacourse KA, Swanberg LJ, Gillespie PJ, Rehfeld JF, Saunders TL and Samuelson LC. Pancreatic function in CCK-deficient mice: adaptation to dietary protein does not require CCK. Am J Physiol 276(5 Pt 1): G1302-1309,1999. PMID: 10330022.
- Laplante M and Sabatini DM. mTOR signaling in growth control and disease. Cell 149(2): 274-293,2012. PMID: 22500797.
- Lazarus SS and Volk BW. Electron Microscopy and Histochemistry of Rabbit Pancreas in Protein Malnutrition (Experimental Kwashiorkor). Am J Pathol 44: 95-111,1964. PMID: 14103205.
- Lhoste EF, Aprahamian M, Balboni G and Damge C. Evidence for a direct trophic effect of bombesin on the mouse pancreas: in vivo and cell culture studies. Regul Pept 24(1): 45-54,1989. PMID: 2472647.
- Li Y, Corradetti MN, Inoki K and Guan KL. TSC2: filling the GAP in the mTOR signaling pathway. Trends Biochem Sci 29(1): 32-38,2004. PMID: 14729330.
- Liddle RA. Regulation of cholecystokinin secretion by intraluminal releasing factors. Am J Physiol 269(3 Pt 1): G319-327,1995. PMID: 7573441.
- Liddle RA, Goldfine ID and Williams JA. Bioassay of plasma cholecystokinin in rats: effects of food, trypsin inhibitor, and alcohol. Gastroenterology 87(3): 542-549,1984. PMID: 6204904.
- Liddle RA, Green GM, Conrad CK and Williams JA. Proteins but not amino acids, carbohydrates, or fats stimulate cholecystokinin secretion in the rat. Am J Physiol 251(2 Pt 1): G243-248,1986. PMID: 3740265.
- Liddle RA, Toskes PP, Horrow J, Ghali J, Dachman A and Stong D. Lack of trophic pancreatic effects in humans with long-term administration of ximelagatran. Pancreas 32(2): 205-210,2006. PMID: 16552342.
- Liener IE, Goodale RL, Deshmukh A, Satterberg TL, Ward G, DiPietro CM, et al. Effect of a trypsin inhibitor from soybeans (Bowman-Birk) on the secretory activity of the human pancreas. Gastroenterology 94(2): 419-427,1988. PMID: 2446949.
- Lim S, Bae JH, Chun EJ, Kim H, Kim SY, Kim KM, et al. Differences in pancreatic volume, fat content, and fat density measured by multidetector-row computed tomography according to the duration of diabetes. Acta Diabetol 51(5): 739-748,2014. PMID: 24671510.
- Lindberg K, Ronn SG, Tornehave D, Richter H, Hansen JA, Romer J, et al. Regulation of pancreatic beta-cell mass and proliferation by SOCS-3. J Mol Endocrinol 35(2): 231-243,2005. PMID: 16216905.
- Logsdon C. Role of Cholecystokinin in Physiologic and Pathophysiologic Growth of the Pancreas. Gastrointestinal Endocrinology. G Greeley. Totowa, NJ, Humana Press: pp 393-422, 1999.
- Logsdon CD. Stimulation of pancreatic acinar cell growth by CCK, epidermal growth factor, and insulin in vitro. Am J Physiol 251(4 Pt 1): G487-494,1986. PMID: 3020992.
- Logsdon CD, Keyes L and Beauchamp RD. Transforming growth factor-beta (TGF-beta 1) inhibits pancreatic acinar cell growth. Am J Physiol 262(2 Pt 1): G364-368,1992. PMID: 1539670.
- Logsdon CD and Williams JA. Pancreatic acinar cells in monolayer culture: direct trophic effects of caerulein in vitro. Am J Physiol 250(4 Pt 1): G440-447,1986. PMID: 2421585.
- Low-Beer TS, Harvey RF, Davies ER and Read AF. Abnormalities of serum cholecystokinin and gallbladder emptying in celiac disease. N Engl J Med 292(18): 961-963,1975. PMID: 1117928.
- Lu L and Logsdon CD. CCK, bombesin, and carbachol stimulate c-fos, c-jun, and c-myc oncogene expression in rat pancreatic acini. Am J Physiol 263(3 Pt 1): G327-332,1992. PMID: 1415544.
- Lyman RL and Lepkovsky S. The effect of raw soybean meal and trypsin inhibitor diets on pancreatic enzyme secretion in the rat. J Nutr 62(2): 269-284,1957. PMID: 13429425.
- Macauley M, Percival K, Thelwall PE, Hollingsworth KG and Taylor R. Altered volume, morphology and composition of the pancreas in type 2 diabetes. PLoS One 10(5): e0126825,2015. PMID: 25950180.
- Mainz DL, Parks NM and Webster PD, 3rd. Effect of fasting and refeeding on pancreatic DNA synthesis and content. Proc Soc Exp Biol Med 156(2): 340-344,1977. PMID: 928405.
- McCue MD. Starvation physiology: reviewing the different strategies animals use to survive a common challenge. Comp Biochem Physiol A Mol Integr Physiol 156(1): 1-18,2010. PMID: 20060056.
- McLaughlin CL, Baile CA and Peikin SR. Hyperphagia during lactation: satiety response to CCK and growth of the pancreas. Am J Physiol 244(1): E61-65,1983. PMID: 6185005.
- Melmed RN and Bouchier IA. A further physiological role for naturally occurring trypsin inhibitors: the evidence for a trophic stimulant of the pancreatic acinar cell. Gut 10(12): 973-979,1969. PMID: 5391956.
- Meloche S and Pouyssegur J. The ERK1/2 mitogen-activated protein kinase pathway as a master regulator of the G1- to S-phase transition. Oncogene 26(22): 3227-3239,2007. PMID: 17496918.
- Messier B and Leblond CP. Cell proliferation and migration as revealed by radioautography after injection of thymidine-H3 into male rats and mice. Am J Anat 106: 247-285,1960. PMID: 13769795.
- Morisset J. Hormonal control of pancreatic growth during fetal, neonatal and adult life. Adv Med Sci 53(2): 99-118,2008. PMID: 18930876.
- Morisset J, Guan D, Jurkowska G, Rivard N and Green GM. Endogenous cholecystokinin, the major factor responsible for dietary protein-induced pancreatic growth. Pancreas 7(5): 522-529,1992. PMID: 1381095.
- Morisset J, Jolicoeur L, Caussignac Y and Solomon TE. Trophic effects of chronic bethanechol on pancreas, stomach, and duodenum in rats. Can J Physiol Pharmacol 60(7): 871-876,1982. PMID: 7127210.
- Morisset JA and Webster PD. Effects of fasting and feeding on protein synthesis by the rat pancreas. J Clin Invest 51(1): 1-8,1972. PMID: 5007051.
- Muller MK, Goebell H, Alfen R, Ehlers J, Jager M and Plumpe H. Effects of camostat, a synthetic protease inhibitor, on endocrine and exocrine pancreas of the rat. J Nutr 118(5): 645-650,1988. PMID: 2452864.
- Muller R, Laucke R, Trimper B and Cossel L. Pancreatic cell proliferation in normal rats studied by in vivo autoradiography with 3H-thymidine. Virchows Arch B Cell Pathol Incl Mol Pathol 59(3): 133-136,1990. PMID: 1980759.
- Nagathihalli NS, Castellanos JA, Shi C, Beesetty Y, Reyzer ML, Caprioli R, et al. Signal Transducer and Activator of Transcription 3, Mediated Remodeling of the Tumor Microenvironment Results in Enhanced Tumor Drug Delivery in a Mouse Model of Pancreatic Cancer. Gastroenterology 149(7): 1932-1943 e1939,2015. PMID: 26255562.
- Nagy I, Pap A and Varro V. Time-course of changes in pancreatic size and enzyme composition in rats during starvation. Int J Pancreatol 5(1): 35-45,1989. PMID: 2473146.
- Nagy I, Takacs T and Mohacsi G. One-year treatment with cholecystokinin-octapeptide and secretin: effects on pancreatic trophism in the rat. Pharmacol Res 36(1): 77-85,1997. PMID: 9368919.
- Nakamura Y, Higuchi S and Maruyama K. Pancreatic volume associated with endocrine and exocrine function of the pancreas among Japanese alcoholics. Pancreatology 5(4-5): 422-431,2005. PMID: 15985767.
- Nevalainen TJ and Janigan DT. Degeneration of mouse pancreatic acinar cells during fasting. Virchows Arch B Cell Pathol 15(2): 107-118,1974. PMID: 4211904.
- Niederau C, Liddle RA, Williams JA and Grendell JH. Pancreatic growth: interaction of exogenous cholecystokinin, a protease inhibitor, and a cholecystokinin receptor antagonist in mice. Gut 28 Suppl: 63-69,1987. PMID: 2446964.
- Niederau C, Niederau M, Klonowski H, Luthen R and Ferrell LD. Effect of hypergastrinemia and blockade of gastrin-receptors on pancreatic growth in the mouse. Hepatogastroenterology 42(4): 423-431,1995. PMID: 8586382.
- Oates PS and Morgan RG. Cell proliferation in the exocrine pancreas during development. J Anat 167: 235-241,1989. PMID: 2630538.
- Oates PS and Morgan RG. Pancreatic growth and cell turnover in the rat fed raw soya flour. Am J Pathol 108(2): 217-224,1982. PMID: 6086052.
- Oates PS and Morgan RG. Random or selective cell death during pancreatic involution following withdrawal of raw soya flour feeding in the rat. Pathology 18(2): 234-236,1986. PMID: 2429247.
- Oates PS, Morgan RG and Light AM. Cell death (apoptosis) during pancreatic involution after raw soya flour feeding in the rat. Am J Physiol 250(1 Pt 1): G9-14,1986. PMID: 2417499.
- Ohlsson B, Borg K, Rehfeld JF, Axelson J and Sundler F. The method of administration of cholecystokinin determines the effects evoked in the pancreas. Pancreas 23(1): 94-101,2001. PMID: 11451154.
- Paredes JL, Orabi AI, Ahmad T, Benbourenane I, Tobita K, Tadros S, et al. A non-invasive method of quantifying pancreatic volume in mice using micro-MRI. PLoS One 9(3): e92263,2014. PMID: 24642611.
- Penzo-Mendez AI and Stanger BZ. Organ-Size Regulation in Mammals. Cold Spring Harb Perspect Biol 7(9): a019240,2015. PMID: 26187729.
- Pfeiffer CJ, Chernenko GA, Kohli Y and Barrowman JA. Trophic effects of cholecystokinin octapeptide on the pancreas of the Syrian hamster. Can J Physiol Pharmacol 60(3): 358-362,1982. PMID: 6176305.
- Pierre JF, Neuman JC, Brill AL, Brar HK, Thompson MF, Cadena MT, et al. The gastrin-releasing peptide analog bombesin preserves exocrine and endocrine pancreas morphology and function during parenteral nutrition. Am J Physiol Gastrointest Liver Physiol 309(6): G431-442,2015. PMID: 26185331.
- Pitchumoni CS. Pancreas in primary malnutrition disorders. Am J Clin Nutr 26(3): 374-379,1973. PMID: 4632067.
- Povoski SP, Zhou W, Longnecker DS, Jensen RT, Mantey SA and Bell RH, Jr. Stimulation of in vivo pancreatic growth in the rat is mediated specifically by way of cholecystokinin-A receptors. Gastroenterology 107(4): 1135-1146,1994. PMID: 7523219.
- Racela AS, Jr., Grady HJ, Higginson J and Svoboda DJ. Protein deficiency in rhesus monkeys. Am J Pathol 49(3): 419-443,1966. PMID: 4958516.
- Regan PT and DiMagno EP. Exocrine pancreatic insufficiency in celiac sprue: a cause of treatment failure. Gastroenterology 78(3): 484-487,1980. PMID: 7351287.
- Richter BD and Schneeman BO. Pancreatic response to long-term feeding of soy protein isolate, casein or egg white in rats. J Nutr 117(2): 247-252,1987. PMID: 3559741.
- Rivard N, Guan D, Maouyo D, Grondin G, Berube FL and Morisset J. Endogenous cholecystokinin release responsible for pancreatic growth observed after pancreatic juice diversion. Endocrinology 129(6): 2867-2874,1991. PMID: 1954873.
- Rozengurt E. Early signals in the mitogenic response. Science 234(4773): 161-166,1986. PMID: 3018928.
- Saisho Y, Butler AE, Meier JJ, Monchamp T, Allen-Auerbach M, Rizza RA, et al. Pancreas volumes in humans from birth to age one hundred taking into account sex, obesity, and presence of type-2 diabetes. Clin Anat 20(8): 933-942,2007. PMID: 17879305.
- Sans MD, Sabbatini ME, Ernst SA, D'Alecy LG, Nishijima I and Williams JA. Secretin is not necessary for exocrine pancreatic development and growth in mice. Am J Physiol Gastrointest Liver Physiol 301(5): G791-798,2011. PMID: 21852360.
- Sans MD, Tashiro M, Vogel NL, Kimball SR, D'Alecy LG and Williams JA. Leucine activates pancreatic translational machinery in rats and mice through mTOR independently of CCK and insulin. J Nutr 136(7): 1792-1799,2006. PMID: 16772439.
- Sans MD, Xie Q and Williams JA. Regulation of translation elongation and phosphorylation of eEF2 in rat pancreatic acini. Biochem Biophys Res Commun 319(1): 144-151,2004. PMID: 15158453.
- Sarfati PD, Genik P and Morisset J. Caerulein and secretin induced pancreatic growth: a possible control by endogenous pancreatic somatostatin. Regul Pept 11(3): 261-273,1985. PMID: 2865774.
- Satake K, Mukai R, Kato Y and Umeyama K. Effects of cerulein on the normal pancreas and on experimental pancreatic carcinoma in the Syrian golden hamster. Pancreas 1(3): 246-253,1986. PMID: 2437568.
- Sato N, Suzuki S, Kanai S, Ohta M, Jimi A, Noda T, et al. Different effects of oral administration of synthetic trypsin inhibitor on the pancreas between cholecystokinin-A receptor gene knockout mice and wild type mice. Jpn J Pharmacol 89(3): 290-295,2002. PMID: 12184735.
- Sato T, Ito K, Tamada T, Sone T, Noda Y, Higaki A, et al. Age-related changes in normal adult pancreas: MR imaging evaluation. Eur J Radiol 81(9): 2093-2098,2012. PMID: 21906894.
- Schaefer J. The normal weight of the pancreas in the adult human being: A biometric study. The Anatomical Record 32(2): 119-132,1926. PMID.
- Schmidt WE, Choudhury AR, Siegel EG, Loser C, Conlon JM, Folsch UR, et al. CCK-antagonist L-364,718: influence on rat pancreatic growth induced by caerulein and bombesin-like peptides. Regul Pept 24(1): 67-79,1989. PMID: 2544930.
- Secor SM and Diamond J. A vertebrate model of extreme physiological regulation. Nature 395(6703): 659-662,1998. PMID: 9790187.
- Sengupta S, Peterson TR and Sabatini DM. Regulation of the mTOR complex 1 pathway by nutrients, growth factors, and stress. Mol Cell 40(2): 310-322,2010. PMID: 20965424.
- Sesso A, Abrahamsohn PA and Tsanaclis A. Acinar cell proliferation in the rat pancreas during early postnatal growth. Acta Physiol Lat Am 23(1): 37-50,1973. PMID: 4713544.
- Smith JP, Kramer ST and Solomon TE. CCK stimulates growth of six human pancreatic cancer cell lines in serum-free medium. Regul Pept 32(3): 341-349,1991. PMID: 1866474.
- Snook JT. Effect of age and long-term diet on exocrine pancreas of the rat. Am J Physiol 228(1): 262-268,1975. PMID: 1147019.
- Snook JT. Effect of diet on development of exocrine pancreas of the neonatal rat. Am J Physiol 221(5): 1388-1391,1971. PMID: 5124284.
- Solomon TE, Morisset J, Wood JG and Bussjaeger LJ. Additive interaction of pentagastrin and secretin on pancreatic growth in rats. Gastroenterology 92(2): 429-435,1987. PMID: 3792779.
- Solomon TE, Petersen H, Elashoff J and Grossman MI. Interaction of caerulein and secretin on pancreatic size and composition in rat. Am J Physiol 235(6): E714-719,1978. PMID: 736132.
- Solomon TE, Vanier M and Morisset J. Cell site and time course of DNA synthesis in pancreas after caerulein and secretin. Am J Physiol 245(1): G99-105,1983. PMID: 6191579.
- Stanger BZ. Organ size determination and the limits of regulation. Cell Cycle 7(3): 318-324,2008. PMID: 18235243.
- Stanger BZ, Tanaka AJ and Melton DA. Organ size is limited by the number of embryonic progenitor cells in the pancreas but not the liver. Nature 445(7130): 886-891,2007. PMID: 17259975.
- Stewart AF, Hussain MA, Garcia-Ocana A, Vasavada RC, Bhushan A, Bernal-Mizrachi E, et al. Human beta-cell proliferation and intracellular signaling: part 3. Diabetes 64(6): 1872-1885,2015. PMID: 25999530.
- Szczepaniak EW, Malliaras K, Nelson MD and Szczepaniak LS. Measurement of pancreatic volume by abdominal MRI: a validation study. PLoS One 8(2): e55991,2013. PMID: 23418491.
- Tashiro M, Dabrowski A, Guo L, Sans MD and Williams JA. Calcineurin-dependent and calcineurin-independent signal transduction pathways activated as part of pancreatic growth. Pancreas 32(3): 314-320,2006. PMID: 16628088.
- Tashiro M, Samuelson LC, Liddle RA and Williams JA. Calcineurin mediates pancreatic growth in protease inhibitor-treated mice. Am J Physiol Gastrointest Liver Physiol 286(5): G784-790,2004. PMID: 14684381.
- Trulsson LM, Svanvik J, Permert J and Gasslander T. Cholecystokinin octapeptide induces both proliferation and apoptosis in the rat pancreas. Regul Pept 98(1-2): 41-48,2001. PMID: 11179777.
- Watanabe H, Saito H, Rychahou PG, Uchida T and Evers BM. Aging is associated with decreased pancreatic acinar cell regeneration and phosphatidylinositol 3-kinase/Akt activation. Gastroenterology 128(5): 1391-1404,2005. PMID: 15887120.
- Watanapa P, Efa EF, Beardshall K, Calam J, Sarraf CE, Alison MR, et al. Inhibitory effect of a cholecystokinin antagonist on the proliferative response of the pancreas to pancreatobiliary diversion. Gut 32(9): 1049-1054,1991. PMID: 1916490.
- Webb SR, Dore BA and Grogan WM. Cell cycle analysis of the postnatal mouse pancreas. Biol Neonate 42(1-2): 73-78,1982. PMID: 7126713.
- Webster PD, Singh M, Tucker PC and Black O. Effects of fasting and feeding on the pancreas. Gastroenterology 62(4): 600-605,1972. PMID: 5020872.
- Weisblum B, Herman L and Fitzgerald PJ. Changes in pancreatic acinar cells during protein deprivation. J Cell Biol 12: 313-327,1962. PMID: 14005814.
- Weiss JM. The ergastoplasm; its fine structure and relation to protein synthesis as studied with the electron microscope in the pancreas of the Swiss albino mouse. J Exp Med 98(6): 607-618,1953. PMID: 13109112.
- Wenzel G, Stocker E and Heine WD. [Cell proliferation in pancreatic acinar epithelia of rats. Autoradiographic studies with 3 H-thymidine]. Virchows Arch B Cell Pathol 10(2): 118-126,1972. PMID: 4622850.
- Williams AJ, Chau W, Callaway MP and Dayan CM. Magnetic resonance imaging: a reliable method for measuring pancreatic volume in Type 1 diabetes. Diabet Med 24(1): 35-40,2007. PMID: 17227322.
- Williams CD. A nutritional disease of childhood associated with a maize diet. Arch Dis Child 8(48): 423-433,1933. PMID: 21031941.
- Williams JA. Intracellular signaling mechanisms activated by cholecystokinin-regulating synthesis and secretion of digestive enzymes in pancreatic acinar cells. Annu Rev Physiol 63: 77-97,2001. PMID: 11181949.
- Wisner JR, Jr., McLaughlin RE, Rich KA, Ozawa S and Renner IG. Effects of L-364,718, a new cholecystokinin receptor antagonist, on camostate-induced growth of the rat pancreas. Gastroenterology 94(1): 109-113,1988. PMID: 3335282.
- Wullschleger S, Loewith R and Hall MN. TOR signaling in growth and metabolism. Cell 124(3): 471-484,2006. PMID: 16469695.
- Xie J, El Sayed NM, Qi C, Zhao X, Moore CE and Herbert TP. Exendin-4 stimulates islet cell replication via the IGF1 receptor activation of mTORC1/S6K1. J Mol Endocrinol 53(1): 105-115,2014. PMID: 24994913.
- Yamamoto M, Otani M, Jia DM, Fukumitsu K, Yoshikawa H, Akiyama T, et al. Differential mechanism and site of action of CCK on the pancreatic secretion and growth in rats. Am J Physiol Gastrointest Liver Physiol 285(4): G681-687,2003. PMID: 12801885.
- Yen JT, Jensen AH and Simon J. Effect of dietary raw soybean and soybean trypsin inhibitor on trypsin and chymotrypsin activities in the pancreas and in small intestinal juice of growing swine. J Nutr 107(1): 156-165,1977. PMID: 556760.
- Zeng LQ, Li FJ, Li XM, Cao ZD, Fu SJ and Zhang YG. The effects of starvation on digestive tract function and structure in juvenile southern catfish (Silurus meridionalis Chen). Comp Biochem Physiol A Mol Integr Physiol 162(3): 200-211,2012. PMID: 22405802.
- Zucker KA, Adrian TE, Bilchik AJ and Modlin IM. Effects of the CCK receptor antagonist L364,718 on pancreatic growth in adult and developing animals. Am J Physiol 257(4 Pt 1): G511-516,1989. PMID: 2478030.