
Methods Type:
Entry Version:
Citation:
Pancreapedia: Exocrine Pancreas Knowledge Base, DOI: 10.3998/panc.2011.6
| Attachment | Size |
|---|---|
| 765.6 KB |
In this chapter we will describe the techniques that can be used to visualise the endoplasmic reticulum (ER) and associated structures in live pancreatic acinar cells.
The ER of pancreatic acinar cells occupies a large portion of the cytoplasm and is densely packed into the basolateral region, with strands threaded into the apical, granular area (4, 10, 12, 19). In this cell type, the ER is responsible for the synthesis of large quantities of digestive enzymes and enzyme precursors (5, 26) and also acts as the principle intracellular calcium (Ca2+) store (2, 15, 18, 19). Junctions with mitochondria (7, 13, 20) and the plasma membrane (14) also enable the ER to affect cellular metabolism and regulate Ca2+ influx. The distribution of ER in pancreatic acinar cells therefore, serves as a useful intracellular landmark that can be correlated to the localisation of other cellular organelles, proteins and signalling events.
Techniques to visualise the ER and associated organelles include the use of cell permeable fluorescent dyes and the exogenous expression of fluorescently tagged ER-resident proteins. Fluorescent dyes are invaluable tools as they can reveal the localisation of intracellular organelles in live cells with minimal disruption (22, 27). ER-Tracker™ Blue-White DPX (ER-Tracker™ B-W) is a cell permeable dye that selectively labels the ER through an unknown mechanism and fluoresces upon excitation with ultraviolet or violet light (6). BODIPY® FL thapsigargin (BODIPY® FL TG) also reveals the ER in living cells. This fluorescent dye is a conjugate of thapsigargin, a drug that binds the sarco/endoplasmic reticulum Ca2+-ATPases (SERCAs) - pumps located in the ER membrane (23). Alternatively, the distribution of the ER can also be revealed by the exogenous expression of fluorescently tagged ER-resident proteins, such as STIM1 (14) or Sec61 (11). Freshly isolated pancreatic acinar cells are notoriously difficult to transfect, but several groups have found that use of an adenovirus vector to deliver constructs into isolated acinar cells allows effective expression of exogenous proteins (16, 17).
Fluorescent dyes can also be used to correlate the distribution of the ER with other cellular structures. FM® 4-64 is a non-cell permeable lipophilic dye that becomes fluorescent upon binding to the outer leaflet of the plasma membrane (3, 21). Tetramethyl rhodamine methyl ester (TMRM) is a cell permeable fluorescent probe with a delocalised positive charge, which accumulates in mitochondria due to the mitochondrial membrane potential (8, 9). Both dyes can be used in conjunction with ER markers to visualise the distribution of the ER relative to the plasma membrane or mitochondria.
This chapter describes the materials and methods required to visualise the ER in live pancreatic acinar cells, and provides examples of how the relative positioning of the ER to other organelles (including the regions of close appositions) can be imaged.
1. Materials
1.1 Isolation of pancreatic acinar cells
- Pancreata are isolated from 6-8 week old male CD-1 mice.
- Extracellular solution – Na+-HEPES based solution (containing in mM: 140 NaCl, 4.7 KCl, 10 HEPES, 1 MgCl2, 10 glucose, 1 CaCl2), adjusted to pH 7.3 using NaOH.
- Purified collagenase (Worthington) stored at -20oC in 1 ml aliquots at 200 units/ml.
- Polycarbonate tubes (Sarstedt).
- Tabletop centrifuge.
- 70 µM cell strainer (BD Biosciences).
- Cover slips or poly-L-lysine coated glass bottom 35 mm culture dishes (MatTek Corp, Ashland, MA, USA).
1.2 Fluorescent dyes
- BODIPY® FL thapsigargin (Molecular Probes) is dissolved in DMSO to give a stock concentration of 1 mM and stored at -20oC.
- ER-Tracker™ Blue-White DPX (Invitrogen) is stored at -20oC.
- FM® 4-64 (Invitrogen) is dissolved in distilled water to give a stock concentration of 1 mM and is stored at -20oC.
- TMRM (Invitrogen) is dissolved in water to give a 50 mM solution, then further diluted in water to give a stock solution of 50 µM and stored at -20oC.
1.3 Transfection of acinar cells using an adenovirus vector
- Acinar cell culture media – Extracellular solution supplemented with MEM amino acids, 292 µg/ml L-glutamine, 100 units/ml penicillin, 100 µg/ml streptomycin, 1 mg/ml trypsin inhibitor, 2.5 pM CCK. Adjusted to pH 7.4 using NaOH and sterile filtered using a Minisart single use filter unit (Sartorius Stedim biotech, Aubagne, France).
- STIM1-EYFP adenovirus (DNA construct produced by Lee Haynes and inserted into a replication-deficient adenovirus by Vector Biolabs, Philadelphia, PA, USA).
- A standard humidified cell culture incubator (Wolf Laboratories, York, UK).
1.4 Depletion of ER Ca2+ to reveal STIM1 at ER-plasma membrane junctions
- Thapsigargin (Calbiochem) is dissolved in DMSO to a stock concentration of 10 mM and stored at -20oC.
2. Methods
2.1 Pancreatic acinar cell isolation
- Mice are humanely killed in accordance with the Animals (Scientific Procedures) Act 1986. The pancreas is dissected out rapidly and washed in extracellular solution, injected with 1 ml (200 units) of collagenase solution and transferred in a 1.5 ml tube to a shaking water bath at 37oC and 120 rpm.
- After 15 minutes, the collagenase solution is removed and the tissue is disrupted in fresh extracellular solution by pipetting through a 1 ml pipette tip with 5 mm of the tip removed. The cloudy supernatant is transferred to a 15 ml polycarbonate tube and the cycle repeated several times.
- The supernatant is then spun at 1000 rpm (260 x g) for 1 minute in a tabletop centrifuge and the resulting pellet resuspended in extracellular solution. Cells are then filtered through a 70 µm cell strainer and the filtrate washed twice by centrifugation. The resulting pellet (composed of isolated cells and small clusters) is then resuspended in extracellular solution and plated onto cover slips or 35 mm glass bottom culture dishes.
2.2 Loading of fluorescent dyes
- BODIPY® FL TG – Cells are incubated with 1 µM BODIPY® FL TG diluted in extracellular solution for 30 minutes at 35oC, excess dye is then washed off using extracellular solution (see Figure 1A and Notes 1 & 2). Staining is also possible at room temperature.
- ER-Tracker™ B-W – Cells are incubated with 1 µM ER Tracker™ B-W for 30 minutes at 35oC, and then washed with extracellular solution (see Figures 1B & C). Staining is also possible at room temperature.
- TMRM – Cells are incubated with 25-50 nM TMRM for 10 minutes at room temperature, and washed with extracellular solution (see Figure 2C).
- FM® 4-64 – Cells are treated with 5 µM FM® 4-64 for approximately 5 minutes before imaging (see Figures 1A & B and Note 3). Staining is reversible and can be removed by washing cells with extracellular solution.
2.3 Transfection of acinar cells using an adenovirus vector
- Freshly isolated cells are resuspended in acinar cell culture media and approximately 100 µl of the cell suspension is seeded onto 35 mm glass bottom dishes. Cells are allowed to attach for 5 minutes before the addition of 2 ml acinar cell culture media.
- Cells are then transfected with an adenovirus vector carrying the STIM1-EYFP construct at approximately 1-5x107 PFU/ml. The required volume of virus is diluted in 100 µl acinar cell culture media and added in a drop-wise fashion to the cells.
- Cells are kept in a standard humidified cell culture incubator at 35oC, 5% CO2 and 95% humidity for approximately 12-14 hours before imaging (see Figures 1 – 3 and Note 4 – Note 6).
2.4 Depletion of ER Ca2+ to reveal STIM1 at ER-plasma membrane junctions
- Cells are incubated with 2 µM thapsigargin diluted in extracellular solution for 10-20 minutes before imaging.
2.5 Imaging
- Cells are imaged using an inverted laser scanning confocal microscope. The images shown in Figure 1-3 are taken using a 63x oil immersion objective with the pinhole set to 1 Airy unit. The images were taken with 12 bit resolution. Pixel size was usually set between 100 and 150 nm. Images were acquired using Leica Confocal Software 2.61. For quantification of fluorescence changes (see Fig 2A) the regions of interest were placed over selected cells, the intensity of fluorescence in each region was acquired and then analysed off-line using Origin 8.1 software (OriginLab). Peak excitation and emission spectra for each fluorophore are shown in Table 1.
| Fluorophore | Peak Excitation (excitation laser line; nm) |
Emission (nm) |
| BODIPY® FL thapsigargin | <488 (488) | <495-550 |
| ER Tracker Blue-White | 374 (405) | 430-640 |
| FM 4-64 | 558 (543) | 675-775 |
| TMRM | 549 (543) | 560-600 |
| STIM1-EYFP | 514 (514) | 520-560 |
Table 1: Excitation and emission wavelengths of fluorophores used to visualise the ER in live isolated pancreatic acinar cells. Recommended peak excitation and the optimal emission range are given, with the excitation laser line used in our laboratory shown in brackets.
3. Notes
- The Golgi apparatus has also been reported to contain SERCA pumps (25). Therefore the use of BODIPY® FL TG can also reveal elements of the Golgi network.
- Thapsigargin is known to deplete ER Ca2+ stores and stimulate Ca2+ influx via store-operated Ca2+ entry (24); it is expected that BODIPY® FL TG will have similar effects. Prolonged treatment can therefore lead to acinar cell damage due to Ca2+ overload, so it is advisable to image cells immediately after loading. The specificity of BODIPY® FL TG can be verified by its displacement with a non-conjugated thapsigargin (12).
- FM® 4-64 can undergo endocytosis, which can subsequently lead to non-specific staining of intracellular lipids (3, 21). For best results, imaging should be carried out shortly after loading.
- Incubation at 35oC is preferable, as cells undergo blebbing at more physiological temperatures (37-38oC), particularly when stimulated with Ca2+-releasing agonists.
- Transfecting acinar cells with adenoviruses carrying DNA constructs can result in excessive levels of exogenous protein. We recommend that the time course of protein expression should be determined for each protein and experiment (see Figure 2A for an example).
- It can be advantageous to compare the distribution of ER proteins in both live (using exogenous protein expression) and fixed cells (using antibodies against the endogenous proteins) to highlight any artefacts occurring due to excessive protein expression (see Figure 3).

Figure 1. ER distribution in live pancreatic acinar cells. Isolated mouse pancreatic acinar cells were loaded with (A) BODIPY® FL TG (blue) and the plasma membrane marker FM® 4-64 (yellow), or (B) ER-Tracker™ B-W (magenta) and FM® 4-64 (yellow). (C) ER-Tracker™ B-W (magenta) was loaded into acinar cells transfected with an adenovirus carrying the STIM1-EYFP (green) construct. In all panels, merged images are shown on the right. Scale bars represent 10 µm.


Figure 2. STIM1 expression in live isolated pancreatic acinar cells reveals ER and ER-plasma membrane junctions. (A) Development of fluorescence over time in pancreatic acinar cells transfected with an adenovirus carrying a STIM1-EYFP construct. (B) STIM1 distribution in different optical sections in resting and (thapsigargin-induced) store-depleted acinar cells expressing exogenous STIM1-EYFP (green). TG: thapsigargin. Scale bars represent 10 µm. (C) Distribution of STIM1-EYFP (green) in relation to mitochondria (magenta; shown using TMRM) in store-depleted acinar cells. The panel on the right shows the relative positioning of the two organelles, with arrows indicating STIM1 puncta. STIM1 puncta can often be seen in the immediate vicinity of mitochondria and the plasma membrane. Parts B and C were adapted from (14).

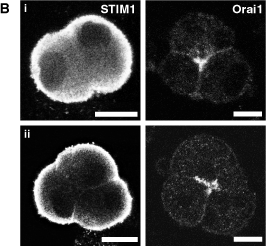
Figure 3. A comparison of STIM1-EYFP and Orai1-mCherry distribution in live and fixed pancreatic acinar cells. (A) Acinar cells transfected with two adenoviruses carrying STIM1-EYFP (green) and Orai1-mCherry (red) constructs in (i) resting and (ii) (thapsigargin-induced) store-depleted conditions. In the merged images (shown in the right panel), yellow punctate structures in the basolateral region highlight ER-plasma membrane junctions. (B) STIM1 and Orai1 distribution in (i) resting and (ii) store-depleted methanol fixed acinar cells, shown using antibodies against endogenous proteins. The Orai1 antibody (14) was produced by Dr. S. Feske. The STIM1 antibody (1) was a gift from Dr. A. Rao. Scale bars represent 10 µm. Figure 3 was adapted from (14). Comparison of figures 3A and 3B illustrates the quality of images attainable in live cells with expressed proteins and fixed cells with immunostaining.
4. Acknowledgements
The work of the group was supported by MRC (UK) grant G0700167, Wellcome Trust grant 080906/Z/06/Z to G.L. and Wellcome Trust grant 086738/Z/08/A to H.D.
5. References
- Barr VA, Bernot KM, Srikanth S, Gwack Y, Balagopalan L, Regan CK, Helman DJ, Sommers CL, Oh-Hora M, Rao A, and Samelson LE. Dynamic movement of the calcium sensor STIM1 and the calcium channel Orai1 in activated T-cells: puncta and distal caps. Mol Biol Cell 19: 2802-2817, 2008. PMID: 18448669
- Berridge MJ, Bootman MD, and Roderick HL. Calcium signalling: dynamics, homeostasis and remodelling. Nat Rev Mol Cell Biol 4: 517-529, 2003. PMID: 12838335
- Betz WJ, Mao F, and Bewick GS. Activity-dependent fluorescent staining and destaining of living vertebrate motor nerve terminals. J Neurosci 12: 363-375, 1992. PMID: 1371312
- Bolender RP. Stereological analysis of the guinea pig pancreas. I. Analytical model and quantitative description of nonstimulated pancreatic exocrine cells. The Journal of Cell Biology 61: 269-287, 1974. PMID: 4363955
- Chen X, Karnovsky A, Sans MD, Andrews PC, and Williams JA. Molecular characterization of the endoplasmic reticulum: Insights from proteomic studies. Proteomics. PMID: 21046616
- Cole L, Davies D, Hyde GJ, and Ashford AE. ER-Tracker dye and BODIPY-brefeldin A differentiate the endoplasmic reticulum and golgi bodies from the tubular-vacuole system in living hyphae of Pisolithus tinctorius. J Microsc 197: 239-249, 2000. PMID: 10692127
- Csordás G, Thomas AP, and Hajnóczky G. Quasi-synaptic calcium signal transmission between endoplasmic reticulum and mitochondria. The EMBO Journal 18: 96-108, 1999. PMID: 9878054
- Duchen MR, Surin A, and Jacobson J. Imaging mitochondrial function in intact cells. Meth Enzymol 361: 353-389, 2003. PMID: 12624920
- Ehrenberg B, Montana V, Wei MD, Wuskell JP, and Loew LM. Membrane potential can be determined in individual cells from the nernstian distribution of cationic dyes. Biophysical Journal 53: 785-794, 1988. PMID: 3390520
- Fogarty KE, Kidd JF, Turner A, Skepper JN, Carmichael J, and Thorn P. Microtubules regulate local Ca2+ spiking in secretory epithelial cells. J Biol Chem 275: 22487-22494, 2000. PMID: 10801885
- Friedman JR, Webster BM, Mastronarde DN, Verhey KJ, and Voeltz GK. ER sliding dynamics and ER-mitochondrial contacts occur on acetylated microtubules. J Cell Biol 190: 363-375. PMID: 20696706
- Gerasimenko OV, Gerasimenko JV, Rizzuto RR, Treiman M, Tepikin AV, and Petersen OH. The distribution of the endoplasmic reticulum in living pancreatic acinar cells. Cell Calcium 32: 261-268, 2002. PMID: 12543088
- Johnson PR, Tepikin AV, and Erdemli G. Role of mitochondria in Ca(2+) homeostasis of mouse pancreatic acinar cells. Cell Calcium 32: 59-69, 2002. PMID: 12161106
- Lur G, Haynes LP, Prior IA, Gerasimenko OV, Feske S, Petersen OH, Burgoyne RD, and Tepikin AV. Ribosome-free terminals of rough ER allow formation of STIM1 puncta and segregation of STIM1 from IP(3) receptors. Curr Biol 19: 1648-1653, 2009. PMID: 19765991
- Nathanson MH, Padfield PJ, O'Sullivan AJ, Burgstahler AD, and Jamieson JD. Mechanism of Ca2+ wave propagation in pancreatic acinar cells. J Biol Chem 267: 18118-18121, 1992. PMID: 1517244
- Nicke B, Tseng MJ, Fenrich M, and Logsdon CD. Adenovirus-mediated gene transfer of RasN17 inhibits specific CCK actions on pancreatic acinar cells. Am J Physiol 276: G499-506, 1999. PMID: 9950825
- Padfield PJ, Elliott AC, and Baldassare JJ. Adenovirus-mediated gene expression in isolated rat pancreatic acini and individual pancreatic acinar cells. Pflugers Arch 436: 782-787, 1998. PMID: 9716713
- Pandol SJ, Schoeffield MS, Fimmel CJ, and Muallem S. The agonist-sensitive calcium pool in the pancreatic acinar cell. Activation of plasma membrane Ca2+ influx mechanism. J Biol Chem 262: 16963-16968, 1987. PMID: 3680283
- Park MK, Petersen OH, and Tepikin AV. The endoplasmic reticulum as one continuous Ca(2+) pool: visualization of rapid Ca(2+) movements and equilibration. The EMBO Journal 19: 5729-5739, 2000. PMID: 11060024
- Rizzuto RR, Pinton P, Carrington W, Fay FS, Fogarty KE, Lifshitz LM, Tuft RA, and Pozzan T. Close contacts with the endoplasmic reticulum as determinants of mitochondrial Ca2+ responses. Science 280: 1763-1766, 1998. PMID: 9624056
- Stenovec M, Solmajer T, Perdih A, Vardjan N, Kreft M, and Zorec R. Distinct labelling of fusion events in rat lactotrophs by FM 1-43 and FM 4-64 is associated with conformational differences. Acta Physiol (Oxf) 191: 35-42, 2007. PMID: 17550407
- Terasaki M, Loew L, Lippincott-Schwartz J, and Zaal K. Fluorescent staining of subcellular organelles: ER, Golgi complex, and mitochondria. Curr Protoc Cell Biol Chapter 4: Unit 4.4, 2001. PMID: 18228364
- Thastrup O, Cullen PJ, Drøbak BK, Hanley MR, and Dawson AP. Thapsigargin, a tumor promoter, discharges intracellular Ca2+ stores by specific inhibition of the endoplasmic reticulum Ca2(+)-ATPase. Proc Natl Acad Sci USA 87: 2466-2470, 1990. PMID: 2138778
- Treiman M, Caspersen C, and Christensen SB. A tool coming of age: thapsigargin as an inhibitor of sarco-endoplasmic reticulum Ca(2+)-ATPases. Trends Pharmacol Sci 19: 131-135, 1998. PMID: 9612087
- Van Baelen K, Dode L, Vanoevelen J, Callewaert G, De Smedt H, Missiaen L, Parys JB, Raeymaekers L, and Wuytack F. The Ca2+/Mn2+ pumps in the Golgi apparatus. Biochim Biophys Acta 1742: 103-112, 2004. PMID: 15590060
- Yule DI. Pancreatic acinar cells: molecular insight from studies of signal-transduction using transgenic animals. Int J Biochem Cell Biol 42: 1757-1761. PMID: 20637894
- Zorov DB, Kobrinsky E, Juhaszova M, and Sollott SJ. Examining intracellular organelle function using fluorescent probes: from animalcules to quantum dots. Circulation Research 95: 239-252, 2004. PMID: 15297386