Citation:
Pancreapedia: Exocrine Pancreas Knowledge Base, DOI: 10.3998/panc.2017.10
| Attachment | Size |
|---|---|
| 273.79 KB |
1. Introduction
The pancreas is a retroperitoneal organ with its function being dictated by the possession of two morphologically distinct tissues, the exocrine and endocrine pancreas. The endocrine pancreas is organized into islets of Langerhans, consisting of five cell subtypes: α, β, δ, ε, and PP cells that secrete glucagon, insulin, somatostatin, ghrelin, and pancreatic polypeptide, respectively. Islet cells make up only 2% of the adult pancreatic mass. The exocrine pancreas, which is composed of acinar and ductal epithelial cells, accounts for nearly 98% of the adult pancreatic mass (12). The structure of the exocrine and endocrine pancreas has been extensively studied due the clinical importance of pancreas-specific diseases such as diabetes, pancreatic cancer and pancreatitis. However, many of these findings have been extrapolated from mice, which remains the most studied animal model in pancreas research. This focus on mouse is due to difficulty in procuring human pancreas. The mouse pancreas contains approximately 1000-5000 islets, whereas the human pancreas contains approximately 1-15 million islets; their size is similar with an upper diameter of 500-700 um in both mice and humans (8). Pancreatic islets are highly vascularized micro-organs with a capillary network that is five to ten times denser than that of the exocrine pancreas (Figure 1) (10). Other structures within the pancreas that appear highly vascularized are the pancreatic ducts which are enveloped by a dense network of vessels that are much denser than in the surrounding acinar tissue (Figure 1) (10). Blood vessel formation plays an essential role in adult tissue function as well as during embryonic development (16).

Figure 1: (A) Whole-mount image of wild-type mouse pancreatic tissue demonstrating the hypervascular microcapillary network of the islet with glucagon in the periphery of the islet. (B) Pancreatic duct (*) running parallel to a vessel (arrow head) and enveloped by a dense network of vessels, almost as dense as the islet, as opposed to the acinar vessels (arrow). (C) A higher magnification to demonstrate the hypervascularity of the islet and pancreatic duct. (D) A 3-dimensional reconstruction of (A). CD31: an endothelial marker, also known as anti-platelet/endothelial cell adhesion molecule-1 (PECAM).
2. History
Over time, the pancreas has evolved from relative obscurity, to possessing the most studied cell in the world, the beta cell. The term pancreas, “all-flesh”, was first coined by Aristotle (384 – 322 B.C.) and later described by the Greek anatomist and surgeon Herophilus (335 B.C. to 280 B.C.) (30). However, this organ was then overlooked for many years, partly because of its location as opposed to other more prominent organs such as the liver or kidneys, but mostly because its function was unknown for many centuries. Galen’s (Claudius Galenus, 129 – 216 A.D.) false dogma that the organ functioned as a mechanical protective cushion for the mesenteric vessels was unchallenged until the seventeenth century (17). In 1642, the main pancreatic duct was discovered by the German anatomist Johann Georg Wirsüng (1589–1643) during an autopsy of a human subject, engraving his finding on a copper plate as a method of reproducing the anatomical illustration (17). Although its function was never known to him, it was eventually named after him in honor of his discovery. Though Wirsüng himself and others did make the observation of occasionally seeing the duct to be double, the discovery of the accessory pancreatic duct as a normal variant is credited to Giovanni Domenico Santorini (1681–1737) (17). The pancreatic duct discovery was a milestone in pancreatic history because it made Galen’s theory obsolete, and led to studies of the pancreas as an exocrine gland. Reignier de DeGraaf (1641–1673) showed that the pancreas was undoubtedly an exocrine gland by cannulating the duct, which abolished the previous theory that the pancreas acted as a cushion of the stomach. Paul Langerhans (1847–1888), a German pathologist, was working on his thesis entitled, “Contributions to the microscopic anatomy of the pancreas” where he discovered discrete areas of ‘clear cells’ within the pancreas. Langerhans documented the histology of the pancreas by injecting the ductal system in the rabbit pancreas with Berlin blue dye, identifying and describing the endocrine cells as “small irregular-polygonal structures with a cytoplasm perfectly shiny”. However, the first to discover that these ‘islands of clear cells’ were well vascularized and communicated with the capillary bed of the exocrine tissue were Wilhelm F. Kuhne and A. S. Lea in 1882 who compared these distinct vascular regions to the renal glomeruli (14). In 1893, the French histologist Edouard Laguesse suggested that these vascular regions be named ‘Islets of Langerhans’, in memory of their original discoverer (14).
Since then, a variety of injection techniques have been used by a variety of authors to demonstrate the islet microvascular anatomy including Berlin blue dye, methylene blue, dyed collagenase, and fluorescent dyes such as Lucifer Yellow or tomato lectin (3, 13, 23, 29). Other methods utilized to visualize the microvascular anatomy included corrosion casting coupled with scanning electron microscopy (6). The latter method allowed beautiful three-dimensional view of the pancreatic and islet vasculature, but was labor intensive. Then came the development of diffusible tracer techniques to estimate blood flow distributions within organs, including the islets (26). Microspheres measuring 10-15 μm introduced through an intra-arterial injection became the standard marker for islet blood flow measurements. The number of microspheres within each organ is proportional to their blood perfusion, quantified by either directly by counting the spheres, or by quantifying a signal (radioactivity, fluorescence, or different colors) (18). However, there was a risk of overestimating blood flow due to shunting, and if the number and size of the spheres exceeded a certain threshold, there would be a risk for emboli and tissue ischemia (18). At the turn of the 21st century, novel imaging techniques were developed to visualize the intricate islet microvascular anatomy in three-dimensions such as whole-mount immunohistochemistry (Figure 1) and real-time in vivo imaging (4, 9, 10, 21, 25). A major challenge now is to quantify the endocrine cell mass in vivo to better monitor beta cell loss in diabetics or after islet transplant. New pancreas specific biomarkers are being sought for this purpose (2).
3. Embryonic Pancreatic Vascu-lature Development
Despite the well-established role that blood vessels play in the development of different organ systems, their role in pancreatic morphogenesis and differentiation remains poorly understood. However, it is clear that blood vessels play different roles during different stages of pancreatic development, with the potent vasculogenic factor, Vascular Endothelial Growth Factor (VEGF) having an especially important function (1). In the developing pancreas, there are abundant VEGFR2-positive endothelial cells throughout the early embryonic mesenchyme (19, 28). Furthermore, VEGF receptor (VEGFR2) inhibition resulted in abnormal epithelial growth and differentiation, whereas Pdx1-driven overexpression of VEGF resulted in pancreatic growth arrest and islet disruption (1, 19, 22). The avascular dorsal pancreatic bud first evaginates at embryonic day 9.5 (E9.5) towards the aorta and subsequently undergoes extensive morphogenesis, giving rise to a highly branched and tubular epithelium with small capillaries being observed within the surrounding mesenchyme of the pancreatic bud epithelium. At E10.5, some blood flow is seen in the capillaries surrounding the pancreatic bud. Then at E11.5-E12.0, the pancreatic epithelium further remodels, forming bulging ‘tips’ with blood vessels becoming intercalated in between them. These observations suggest that the pancreas gets vascularized in the early stages through a combination of epithelial growth into a pre-existing mesenchymal vascular plexus of patent capillaries, as well as peripheral vasculogenesis. (i.e. de novo development of vessels from angioblasts). The latter process is directed by the early expression of VEGF throughout the pancreatic epithelium at E11.5. The developing endocrine islets and ductal trunk exhibit high VEGF expression, whereas the acini at the epithelial tips lack VEGF expression. This explains why both islets and pancreatic ducts appear highly vascular in the post-natal pancreas compared to the surrounding acini (Figure 1). A primitive primary honeycomb-like primary plexus of vessels appears in the surrounding pancreatic mesenchyme at E10.5, which later forms the veins, followed by the formation of a large central vessel, which is formed along the center of the pancreatic bud by E12.5, becoming an artery. By E13.5, the central vessel becomes progressively integrated into the expanding pancreas and by E14.5, the main central artery becomes more established, with the fusion of more vessels progressing from proximal to distal pancreas. Thus, the veins localize to the periphery, and the arteries more centrally within the pancreas (1).
Shah et al. (28) nicely demonstrated in their whole embryo “angiogram”, utilizing a novel in utero intracardiac injection, a relative paucity of blood flow in early pancreatic development (E11.5), with the presence of blood flow correlating with the differentiation of pancreatic epithelial cells in the region of the blood flow. Specifically, during E12.5–E14.0 glucagon-positive cells tended to localize specifically near perfused blood vessels, and by early E14 insulin-positive cells tended to localize near the blood flow. Thus, endothelial cells and blood flow appear to help direct pancreatic differentiation (19, 28).
4. Gross Anatomy of the Pancreas
The pancreas is an endodermally-derived retroperitoneal organ that is formed by the fusion of the ventral and dorsal buds of the foregut. In the human it is divided into the head, neck, body, and the tail, with no clear cut borders between them. However, it is generally accepted that the border between the head/neck and body is marked by the superior mesenteric artery, whereas the border between the body and tail is considered to be the midpoint between the neck and the tip of the tail. The head has an extension known as the uncinate process that hooks beneath the superior mesenteric artery, with the head nestled in the curvature of the duodenum. The body of pancreas sits beneath the stomach, extending horizontally and crossing over the inferior vena cava, abdominal aorta, superior mesenteric artery, and vein. The neck is the thin section of the gland between head and body overlying the superior mesenteric artery. The pancreatic tail abuts the splenic hilum. The total length of the gland is about 15 cm and weighs 50-100 gm (8).
The mouse pancreas is divided into three major parts: the duodenal, splenic and gastric lobes, with the splenic lobe making up over half of the total pancreatic volume. The duodenal lobe is homologous with the head of the human pancreas, whereas the splenic lobe is homologous with the body and tail of the human pancreas, which extends horizontally between the duodenum and spleen. The gastric lobe is the third and smallest part of the mouse pancreas (8).
The distribution of islets in different lobes of the mouse pancreas is heterogeneous and strain dependent. The number of islets per unit volume is highest in the gastric lobe, followed by the duodenal and splenic lobe. In the human pancreas, the density of islets per unit volume is higher in the tail compared to the head and body (8).
5. Macroscopic Vascular Anatomy of the Pancreas
The blood supply of the pancreas is primarily derived from the celiac artery, representing approximately 1% of the total cardiac output (20). The head of the pancreas is supplied by an arterial arcade system formed by the anterior and posterior superior pancreaticoduodenal arteries arising from the gastroduodenal artery of the common hepatic artery and anastomosing with the anterior and posterior inferior pancreaticoduodenal artery, which is the first branch of the superior mesenteric artery. The body and pancreatic tail on the other hand primarily receive their blood supply from the pancreatic branches of the splenic artery, which is the left most branch of the celiac artery, and by the dorsal pancreatic artery that branches off near the origin of either celiac, hepatic, or splenic artery. Its right branch anastomoses with the anterior superior pancreaticoduodenal artery whereas its left branch forms the transverse pancreatic artery that runs along the inferior border of the pancreas and usually anastomoses with the pancreatica magna artery, the largest pancreatic branch of the splenic artery in the middle of the gland (5, 31).
The venous drainage of the pancreas mainly follows the arterial system and drains into the portal vein via either the splenic vein, from the body and tail of the pancreas, or via the superior mesenteric vein from the pancreatic head venous arcade system (8).
6. Microscopic Vascular Anatomy of the Pancreas
The islets of Langerhans, which compromise nearly 2% of the total pancreatic volume, are highly vascularized structures within the pancreas, receiving approximately 10-fold the blood supply of the exocrine tissue by volume, with the intra-islet microcapillary network constituting 7-8% of the total islet volume (15, 27) (Figure 1). The islets are irregular in shape, but mostly circular to oval, scattered individual endocrine cells, not contained within an islet, are found throughout the pancreatic acinar and ductal tissue (Figure 2). The ratio of beta to alpha cells is higher in mice than in humans, with beta cells making up the majority of endocrine cells within an islet (50-70% and 60-80% in humans and mice, respectively). Alpha cells compose 20-40% and 10-20% of endocrine cells in human and mouse islets, respectively (8, 10) (Figure 3). Depending on the size of the islet, 1-5 arterioles enter each islet and divide into a dense fenestrated capillary network (9) (Figure 4). In most rodents, the distribution of endocrine cells within an islet follows a mantle distribution of non-beta cells, including alpha, delta and pancreatic polypeptide with beta cells filling up the core (Figure 3F). This unique distribution appears to have functional implications. When beta cells are intermingled with non-beta cells instead of the mantle-core segregation, the beta-cell function was reduced (11). Bosco et al. (7) demonstrated in their morphological analysis of the human pancreas that the alpha cell distribution was not random. In small human islets (40-60 um in diameter) the typical mantle-core distribution was seen with alpha cells in the periphery and beta cells occupying the core. In larger human islets (>60 um in diameter) it was noted that alpha-cells were found in a mantle, but they also always lined the inner vascular channels, which are the empty spaces found in the middle of the islet, thus always juxtaposed to endothelial cells. Their three-dimensional analysis revealed that islet cells are organized into a trilaminar plate composed of one layer of beta-cells sandwiched between two layers of alpha-cells. The islets are folded onto each other with different degrees of complexity and vessels invaginating the ‘empty spaces’ within the islets (7).
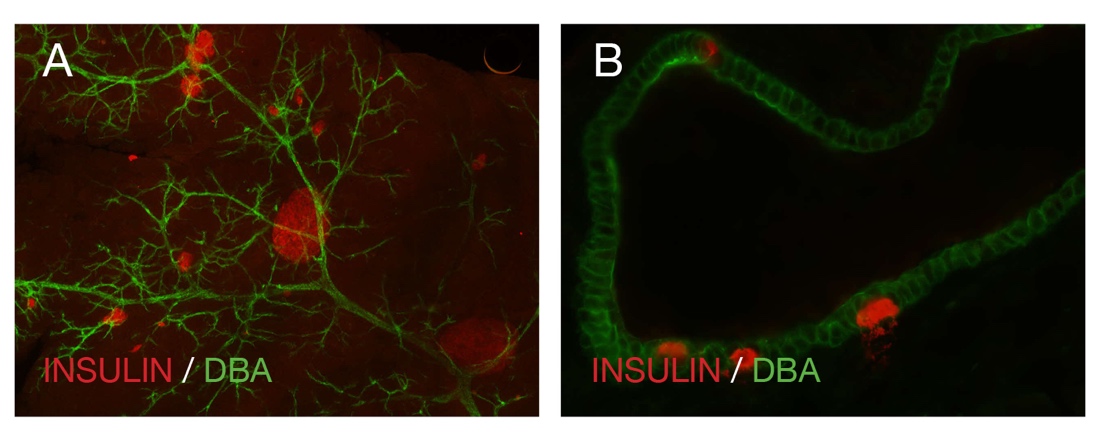
Figure 2: (A) Whole-mount image of wild-type mouse pancreatic tissue demonstrating the pancreatic duct (DBA) and different shapes and sizes of the islets (red). (B) A pancreatic section from a wild-type mouse showing scattered insulin positive endocrine cells (red) in a pancreatic duct (DBA). DBA: dolichos biflorus agglutinin, a ductal marker.

Figure 3: (A) Whole-mount image of a 2 month-old human pancreatic islet. (B-D) Pancreatic section of a 16 year old human pancreatic islet revealing that up to 40% of the islet is composed of alpha cells (C). (E) Section of a 2month old small (<60um) human pancreatic islet demonstrating the mantle-core distribution of alpha and beta cells. (F) Section of a wild-type mouse pancreas revealing the typical mantle distribution of the alpha cells with 80% of the islet composed of insulin secreting beta cells. Pancreatic duct cells are labeled by DBA.

Figure 4: Whole mount confocal microscopy of a mouse islet with arrowheads indicating 5 vessels, 2 arterioles (thin caliber vessel) and 3 venules (larger caliber vessel)
There have been numerous islet vascular perfusion studies conducted and it appears, at least in mice, that there are three microcirculatory flow patterns. Firstly, from center to periphery (i.e. alpha cells are exposed to blood first). Secondly, from center to periphery (i.e. beta cells are exposed to blood first) and lastly, from one pole of the islet to the other. Roughly, 66% of islets have the center-to-periphery pattern (24, 25) (Figure 5).

Figure 5: Three blood flow patterns of the islet microcirculation. (A) From periphery to center. (B) From center to periphery. (C) From one pole to the other
7. References:
- Azizoglu DB, Chong DC, Villasenor A, Magenheim J, Barry DM, Lee S, et al. Vascular development in the vertebrate pancreas. Dev Biol 420(1): 67-78,2016. PMID: 27789228.
- Balhuizen A, Massa S, Mathijs I, Turatsinze JV, De Vos J, Demine S, et al. A nanobody-based tracer targeting DPP6 for non-invasive imaging of human pancreatic endocrine cells. Sci Rep 7(1): 15130,2017. PMID: 29123178.
- Bank HL. Assessment of islet cell viability using fluorescent dyes. Diabetologia 30(10): 812-816,1987. PMID: 3322907.
- Berclaz C, Pache C, Bouwens A, Szlag D, Lopez A, Joosten L, et al. Combined Optical Coherence and Fluorescence Microscopy to assess dynamics and specificity of pancreatic beta-cell tracers. Sci Rep 5: 10385, 2015. PMID: 25988507.
- Bertelli E, Di Gregorio F, Mosca S and Bastianini A. The arterial blood supply of the pancreas: a review. V. The dorsal pancreatic artery. An anatomic review and a radiologic study. Surg Radiol Anat 20(6): 445-452,1998. PMID: 9932331.
- Bonner-Weir S and Orci L. New perspectives on the microvasculature of the islets of Langerhans in the rat. Diabetes 31(10): 883-889,1982. PMID: 6759221.
- Bosco D, Armanet M, Morel P, Niclauss N, Sgroi A, Muller YD, et al. Unique arrangement of alpha- and beta-cells in human islets of Langerhans. Diabetes 59(5): 1202-1210,2010. PMID: 20185817.
- Dolensek J, Rupnik MS and Stozer A. Structural similarities and differences between the human and the mouse pancreas. Islets 7(1): e1024405,2015. PMID: 26030186.
- El-Gohary Y, Sims-Lucas S, Lath N, Tulachan S, Guo P, Xiao X, et al. Three-dimensional analysis of the islet vasculature. Anat Rec (Hoboken) 295(9): 1473-1481,2012. PMID: 22807267.
- El-Gohary Y, Tulachan S, Branca M, Sims-Lucas S, Guo P, Prasadan K, et al. Whole-mount imaging demonstrates hypervascularity of the pancreatic ducts and other pancreatic structures. Anat Rec (Hoboken) 295(3): 465-473,2012. PMID: 22282427.
- Gannon M, Ray MK, Van Zee K, Rausa F, Costa RH and Wright CV. Persistent expression of HNF6 in islet endocrine cells causes disrupted islet architecture and loss of beta cell function. Development 127(13): 2883-2895,2000. PMID: 10851133.
- Gittes GK. Developmental biology of the pancreas: a comprehensive review. Dev Biol 326(1): 4-35,2009. PMID: 19013144.
- Henderson JR and Daniel PM. A comparative study of the portal vessels connecting the endocrine and exocrine pancreas, with a discussion of some functional implications. Q J Exp Physiol Cogn Med Sci 64(4): 267-275,1979. PMID: 118478.
- Henderson JR, Daniel PM and Fraser PA. The pancreas as a single organ: the influence of the endocrine upon the exocrine part of the gland. Gut 22(2): 158-167,1981. PMID: 6111521.
- Henderson JR and Moss MC. A morphometric study of the endocrine and exocrine capillaries of the pancreas. Q J Exp Physiol 70(3): 347-356,1985. PMID: 3898188.
- Herbert SP and Stainier DY. Molecular control of endothelial cell behaviour during blood vessel morphogenesis. Nat Rev Mol Cell Biol 12(9): 551-564,2011. PMID: 21860391.
- Howard JM and Hess W. History of the pancreas : mysteries of a hidden organ In. New York, Kluwer Academic, 2002.
- Jansson L, Barbu A, Bodin B, Drott CJ, Espes D, Gao X, et al. Pancreatic islet blood flow and its measurement. Ups J Med Sci 121(2): 81-95,2016. PMID: 27124642.
- Lammert E, Cleaver O and Melton D. Induction of pancreatic differentiation by signals from blood vessels. Science 294: 564-567,2001. PMID: 11577200.
- Lewis MP, Reber HA and Ashley SW. Pancreatic blood flow and its role in the pathophysiology of pancreatitis. J Surg Res 75(1): 81-89,1998. PMID: 9614861.
- Li G, Wu B, Ward MG, Chong AC, Mukherjee S, Chen S, et al. Multifunctional in vivo imaging of pancreatic islets during diabetes development. J Cell Sci 129(14): 2865-2875,2016. PMID: 27270669.
- Magenheim J, Ilovich O, Lazarus A, Klochendler A, Ziv O, Werman R, et al. Blood vessels restrain pancreas branching, differentiation and growth. Development 138(21): 4743-4752,2011. PMID: 21965615.
- Michaels RL and Sheridan JD. Islets of Langerhans: dye coupling among immunocytochemically distinct cell types. Science 214: 801-803,1981. PMID: 6117129.
- Nyman LR, Ford E, Powers AC and Piston DW. Glucose-dependent blood flow dynamics in murine pancreatic islets in vivo. Am J Physiol Endocrinol Metab 298(4): E807-814,2010. PMID: 20071562.
- Nyman LR, Wells KS, Head WS, McCaughey M, Ford E, Brissova M, et al. Real-time, multidimensional in vivo imaging used to investigate blood flow in mouse pancreatic islets. J Clin Invest 118(11): 3790-3797,2008. PMID: 18846254.
- Prinzen FW and Bassingthwaighte JB. Blood flow distributions by microsphere deposition methods. Cardiovasc Res 45(1): 13-21,2000. PMID: 10728307.
- Saito K, Yaginuma N and Takahashi T. Differential volumetry of A, B and D cells in the pancreatic islets of diabetic and nondiabetic subjects. Tohoku J Exp Med 129(3): 273-283,1979. PMID: 392812.
- Shah SR, Esni F, Jakub A, Paredes J, Lath N, Malek M, et al. Embryonic mouse blood flow and oxygen correlate with early pancreatic differentiation. Dev Biol 349(2): 342-349,2011. PMID: 21050843.
- Shimoda M, Itoh T, Sugimoto K, Iwahashi S, Takita M, Chujo D, et al. Improvement of collagenase distribution with the ductal preservation for human islet isolation. Islets 4(2): 130-137,2012. PMID: 22627378.
- Tsuchiya R and Fujisawa N. On the etymology of "pancreas". Int J Pancreatol 21(3): 269-272,1997. PMID: 9322128.
- Woodburne RT and Olsen LL. The arteries of the pancreas. Anat Rec 111(2): 255-270,1951. PMID: 14894836.