
Entry Version:
Citation:
Pancreapedia: Exocrine Pancreas Knowledge Base, DOI: 10.3998/panc.2016.18
| Attachment | Size |
|---|---|
| 472.45 KB |
1. Introduction
Heat shock proteins (HSPs) are highly conserved proteins, which are expressed in response to stress in all the species. Ritossa in 1962 (31), was first to observe an altered puffing pattern in giant chromosomes of salivary glands in Drosophila busckii after heat shock. On microscopic evaluation, areas of increased transcriptional activity in these giant chromosomes appear swollen and are called puffs. He reported that temperature shock induced well defined variation in the normal puffing patterns, which is observed during the development of Drosophila larvae. After initiation of heat shock, incorporation of 3H-cytidine in puffs started within 3-4 minutes and reached maximum after 10 minutes. In addition to heat shock, this effect could also be reproduced by treatment with 2,4-dinitrophenol (DNP) and sodium salicylate. After the initial report, this important work remained largely ignored till it was rediscovered in 1974. Tissieres et al (38), while studying synthesis of proteins by metabolic incorporation of radioactive precursors, observed a new pattern of radiolabeled protein synthesis following heat shock. These new proteins were termed as “heat shock” proteins. It was observed that heat shock activated synthesis of only a few polypeptides and strongly inhibited the synthesis of most others. Subsequently, it was found that this family of proteins is up-regulated in response to variety of stresses like heat stress, inflammation, ischemia, anoxia and heavy metals and now these are commonly called heat stress proteins (HSPs).
HSPs are subdivided according to their molecular size (from 10kDa till 100 kDa e.g. HSP110, HSP90 (HSP90a, HSP90b. GRP94), HSP70 (HSP70, HSC70, mHSP70 (GRP75), GRP78) (21, 22, 43), HSP60 (6), HSP40 (7, 30), HSP27 (35) and HSP10 (GroES) (13, 15) and contains both constitutively expressed members as well as polypeptides whose expression increases many folds in response to stress.
HSP100: This family is comprised of proteins with MW 104-110kDa and each member has ATPase activity (34). Members of this family include both constitutively expressed and inducible proteins. There are usually two ATP binding domains however a few members have only one ATP binding site. HSP104 is required for inducing thermo-tolerance in yeast and appears to disaggregate insoluble proteins. Proteins of this family are localized to different subcellular compartments (34).
HSP90s: These proteins (MW 82-94kDA) have permanent and abundant expression in eukaryotic cells (14, 20). This family of proteins is involved in regulation of the cytoskeleton dynamics, cell shape and motility. Experiments suggest that HSP90 may be involved in cross linking actin filaments in a Ca2+ dependent fashion and has critical ATPase activity.
HSP70: The HSP70s are ubiquitous and have both constitutive and inducible members (24). These are major molecular chaperones of eukaryotes and are present in all cellular compartments. These have highly conserved nucleotide-binding N-terminal and relatively variable C-terminal domains. Synthesis of inducible members sharply increases in response to stress. The constitutive members are equally important because of their role in protein folding, maturation and proteolysis. HSP72 (HSP70) is best known stress inducible cytoplasmic chaperone, while other stress-inducible chaperones are found in other cell compartments. GRP78 is located in endoplasmic reticulum while GRP75 is found in mitochondria. HSC71 is present in cytoplasm but is constitutively expressed.
HSP60: It is a mitochondrial chaperone but is coded by a nuclear gene (6). It is essential for the folding and assembly of newly imported proteins in mitochondria. HSP60 monomers form a complex arrangement as two stacks of 7 monomers each. This complex binds to unfolded proteins and catalyze their folding in ATP dependent manner.
HSP40: All DnaJ/HSP40 proteins contain J domain through which they bind to HSP70s and can be categorized into three groups depending upon the presence of other domains (30). In humans, 41 DnaJ/HSP40 family members have been identified based upon genome-wide analysis. HSP40s are considered to be co-chaperones for HSP70s because their binding causes increase in ATP hydrolysis activity of HSP70. ATP hydrolysis is essential for activity of HSP70s. In absence of co-chaperone ATP hydrolysis by HSP70s proceeds slowly but in presence of co-chaperone (DnaJ/Hsp40s), it is enhanced several fold thereby accelerating the processing of unfolded polypeptides. DnaJ/Hsp40s also helps in binding of HSP70 to polypeptides, which stabilize their interaction with the substrates. HSP40s activity is affected by the redox status (30).
HSP27: These proteins lack ATPase activity but can be phosphorylated (2, 25, 32). They protect other proteins from thermal inactivation and aggregation. Role of phosphorylation has been studied and it has been suggested that transient phosphorylation might play a role in release of unproductively unfolded proteins from HSP27. It is required for suppression of ASK1 cell death signaling and neuroprotection against ischemic injury (35). Oligomerization of HSP27 is required for chaperon activity, phosphorylation down regulate its chaperone properties by decreasing oligomerization. In response to stress, the expression is increased but it follows a different kinetics compared to phosphorylation which is considered its first response to stress.
Ubiquitin: It is a low molecular weight (~8kDA) heat-inducible protein, which is constitutively expressed in most cell types in mammalian cells and its synthesis increases with heat shock. These have a role in protein degradation. Heat shock response stimulates, protein degradation and increased ubiquitin synthesis facilitates the process (9, 27). The ubiquitin gene constitute a multigene family and three types of mRNA are detectable. The function of ubiquitin is presumed to be labeling of polypeptides designated as substrate for intracellular proteases.
Crystallin: The lens structural proteins, crystallin-αA and -αB are also considered to be HSPs (8). Besides their robust expression in lens, these are also expressed in other tissues. αA-crystallin is expressed in human and mouse pancreas and is involved in modulating activation of AP-1. αA crystallin has also been shown to negatively regulate pancreatic tumorigenesis (8).
2. Expression of Heat Shock Proteins in Pancreatitis
Caerulein- as well as L-arginine-induced models of acute pancreatitis have been used to evaluate heat shock response during development of acute pancreatitis (37, 42). When pancreatic lobules were either stimulated with supramaximal concentrations of caerulein or heat shocked by keeping at 42oC for 60 min, HSP70 mRNA was found to be upregulated (42). The relative expression level of HSP70 was found different in response to induction of pancreatitis and heat shock (42). With caerulein induced pancreatitis, the levels of HSP70 were increased 6-7 fold, but in response to heat shock the expression levels reached up to 20-23 fold. When metabolic labeling of rat acinar cells with 35S-methionine was used to determine hyperthermia-induced expression of proteins, it was found that, overall protein synthesis is reduced by hyperthermia, but proteins with apparent molecular weight of 90, 72, 59, 58 and 30 kilo Daltons were induced. Thermal stress with 42oC temperature led to significantly higher incorporation of radioactivity in at least five acinar proteins of 90, 72, 59, 58 and 30 kilo Dalton apparent molecular weight whereas, incubation at 40oC had no such effect (41). Degree of induction of hyperthermia-induced protein synthesis, however, varied greatly between individual proteins and 72-kilodalton protein showed the greatest and 90kDa the lowest response (8). Four isoforms of HSP70 were expressed in the pancreas in response to thermal stress in vivo.
Heat stress response has been evaluated at 24 h in pancreases of rats after intra-peritoneal injection of L-arginine at 3.0g/kg and 4.5g/kg (37). With induction of arginine pancreatitis there was increase in expression of HSP27, 70, 60 and 90. Lower dose of L-arginine, resulted in highest expression of HSP27 followed by HSP70, 60 and 90 (37). With a higher dose, HSP60 was expressed at higher level followed by HSP27 and HSP70. HSP27 exists as three isoforms (unphosphorylated, mono- and diphosphorylated forms) in rat pancreatic acinar cells. HSP27 phosphorylation was stimulated by CCK in vivo and in vitro and by osmotic stress in vitro. Phosphorylation status of HSP27 was also altered with monophosphorylated form increasing at 6 h and remained elevated up to 120 h (37). Dephosphorylated form was maximum at 12 h and decreased thereafter but was still higher compared to basal level. Injection of nonpathological dose of L-arginine (3g/kg) induced a higher ratio of phosphorylated to unphosphorylated HSP27 protein which indicates that it might be a protective response. Overexpression of HSP27 is known to confer resistance to heat and other stresses. The mechanism of induction of HSP during induction of acute pancreatitis has not been subject of any study specifically. Some of the plausible mechanisms could be (i) decrease in cellular contents of ATP (ii) accumulation of unfolded proteins (iii) ischemia because of compromised microcirculation (29). All of these are capable of triggering stress in the cell and a heat stress response. In the study by Ritossa et al (31)it was found that in addition to heat stress, treatment with dinitrophenol, also altered the puffing pattern. Since dinitrophenol inhibits oxidative phosphorylation and can decrease intracellular ATP levels, this could be a strong signal for triggering stress response. Pancreatitis causing agents like caerulein, bile acids, palmitoleic acid (POA) and palmitoleic acid ethyl ester (POAEE) decrease levels of ATP in pancreas (12, 40).
3. HSP mediated protection from pancreatitis
Based upon the initial observations, which showed that pre-exposure to sub-lethal stress has a protective effect to subsequent lethal exposure, there are several studies, which have investigated expression of heat stress proteins during development of acute pancreatitis (37, 41, 42). In these studies, increased expression of HSPs was observed during acute pancreatitis and it was hypothesized that this is a protective response of acinar cells and pre-induction of this response might protect pancreatic acinar cells from subsequent injury. In pancreatitis, injury is initiated in pancreatic acinar cells which eventually leads to development of acute pancreatitis. Weber et al (1995) (42) studied autoprotective potential of pancreatic acinar cells by exposing rat pancreatic lobules either to caerulein (100nM) stimulation or hyperthermia (42oC for 60 min). The expression of HSP70 and ubiquitin in response to caerulein stimulation and hyperthermia was compared to unexposed pancreatic lobules. The rationale of using pancreatic lobules rather than pancreatic acinar cells was that cells will undergo minimal processing related stress. Increased expression of HSP70 mRNA was observed with caerulein hyperstimulation as well as hyperthermia but there was no effect on ubiquitin expression. This protective effect was further confirmed by induction of mild edematous pancreatitis in rats using caerulein (10 ug/kg/h, i.v.). Increased expression of HSP70 was observed as early as at 4 h and reached maximum by 12 h, however, ubiquitin mRNA didn’t change. It was hypothesized that expression of HSP70 during pancreatitis is a self-defense mechanism of pancreatic acinar cells.
The expression of various HSPs and its correlation with reduced severity of acute pancreatitis was further investigated by Wagner et al (1996) (41). The authors exposed rat pancreatic acini to heat and analyzed protein expression using 35S-methionine labeling and western blots. They observed that after thermal stress (42oC) pancreatic acini increasingly expressed five proteins of 92, 72, 59, 58, and 30kDa, with HSP-70 the most strongly induced 72kDa protein. After whole body hyperthermia (42oC for 20min, using a heat pad and lamp in anesthetized rats), caerulein induced pancreatic organ damage was greatly reduced (41). The degree of protection was correlated to strength of HSP induction. It was proposed that there is a causal relation between hyperthermia-induced HSPs expression and organ protection from subsequent injury to pancreas.
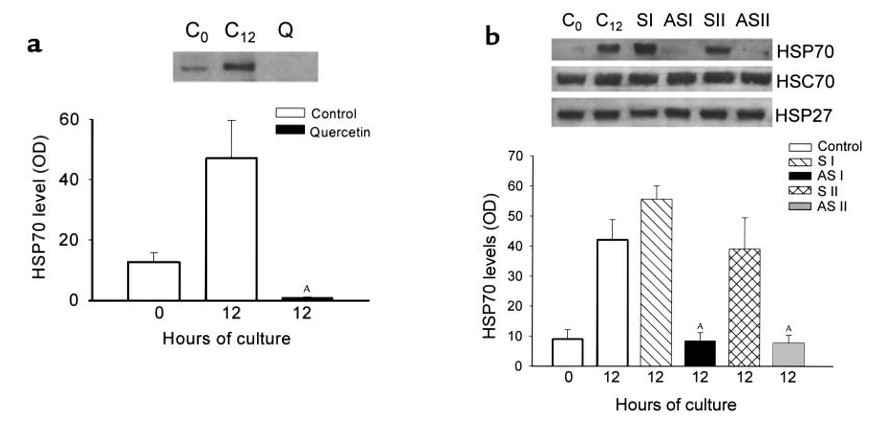
Figure 1. Effect of incubation of pancreas fragments with quercetin or antisense/sense HSP70 oligonucleotides on HSP expression. (a) C0, freshly prepared pancreas fragments; C12, pancreas fragments after 12 hours of culture; Q, pancreas fragments after 12 hours of culture with quercetin (50 μM). HSP70 protein expression was evaluated by Western blotting, and relative optical densities are expressed as mean ± SEM for at least three separate experiments in each group. AP < 0.01 when quercetin-incubated fragments are compared with untreated 12-hour control fragments. (b) C0, freshly prepared pancreas fragments; C12, pancreas fragments after 12 hours of culture; S I, pancreas fragments incubated with sense oligonucleotide S I (1 μM) for 12 hours; AS I, pancreas fragments incubated with 14-mer antisense oligonucleotide AS I (1 μM) for 12 hours; S II, pancreas fragments incubated with sense oligonucleotide S II (1 μM) for 12 hours; AS II, pancreas fragments incubated with 18-mer antisense oligonucleotide AS II (1 μM) for 12 hours. Expression of indicated HSPs was assessed by Western blotting. Relative optical densities of HSP70 bands are expressed as mean ± SEM for at least three separate experiments in each group. AP < 0.01 when antisense oligonucleotide-incubated fragments are compared with untreated 12-hour control fragments. (Reproduced with permission from J Clin Invest) (4)
Because heat stress induced multiple HSPs, although HSP70 (also known as HSP72) was the major protein, it was not possible to predict that the observed protection of pancreatitis is due to HSP70 overexpression. This issue was investigated by Bhagat et al (4) and using in vitro culture of pancreatic lobules it was demonstrated that HSP70 overexpression is actually responsible for the observed protective role of heat stress. In this elegant study, two different approaches, namely selective inhibition of HSP70 overexpression using antisense approach and pharmacological inhibition (Figure 1 & 2) were used to understand the contribution of HSP70 in heat stress induced protection against acute pancreatitis. In the presence of HSP70 anti-sense or pharmacological inhibitor quercetin, the protection against caerulein induced injury was abrogated (Figure 1 & 2). These findings indicated that the protection against caerulein-induced pancreatitis that follows heat stress is mediated by HSP70. The role of HSP70 in protection imparted by heat stress against pancreatic injury was further evaluated in vivo by Bhagat et al (5)(Figure 3). In this study, when antisense oligonucleotide specific to HSP70 was administered prior to heat stress, expression of other stress proteins except HSP70 was observed. The protective effect of pre-induced heat stress to caerulein-induced pancreatitis in these animals was lost indicating that this protective effect is mediated through overexpressed HSP70. But in the group, where sense-oligonucleotide for HSP70 was administered prior to heat stress, HSP70 overexpression was not affected and the protective effect of heat stress to caerulein-induced pancreatitis was maintained.
HSP70 can also be induced by means other than heat stress and administration of sodium arsenite induces overexpression of HSP70 (3). Bhagat et al (3) investigated whether sodium arsenite induced overexpression of HSP70 protects against acute pancreatitis, similar to the HSP70 expression by heat stress. It was observed that sodium arsenite induced HSP70 overexpression provides protection in caerulein (Figure 4 & 5) as well as in L-arginine-induced model of acute pancreatitis. This study also demonstrated that the protective effects of heat stress are not because of non-HSP related heat stress events and is not only limited to mild form of caerulein-induced pancreatitis. Further studies by Frossard et al (10), where HSP70 induced by β-adrenergic stimulation protected against caerulein-induced pancreatitis, support the conclusion that heat stress protein 70 has a protective role in pancreatitis irrespective of mode of induction.
Further evidence of role of HSP70 in modulation of severity of acute pancreatitis came from the clinical studies, where severity of acute pancreatitis was related to HSP70 gene polymorphism. This study revealed that the HSP 70.2 expression was linked to severity of pancreatitis. HSP70-2G allele has been associated with a low HSP70-2 expression and was more prevalent in patients with severe pancreatitis than in patients with mild disease or in healthy population. Conversely patients with “protective” AA genotype are less vulnerable to severe disease and expected to have better prognosis with far fewer complications. In this study, the co-existence of TNF2 and HSP70-2G was detected in 9 of the 49 patients in severe group and 6 of these patients suffered from infected necrosis with multiple organ failure and 1 death (1).
Pancreatic HSP60 has been reported to be induced weakly in response to heat stress but has robust expression as a result of water immersion stress (Lee et al. 2000) (19). Prior induction of HSP60 was also found to be protective against caerulein-induced pancreatitis as evaluated by different markers of severity. Although HSP60 was the main stress protein overexpressed by water immersion, the water immersion can theoretically induce other proteins and pathways which could be responsible for the observed protection, therefore further confirmation of these findings by targeted changes in HSP60 expression is required.

Figure 2. Effect of incubation of pancreas fragments with antisense/sense HSP70 oligonucleotides (1 μM) on trypsinogen activation and cell injury as assessed by LDH release into the medium. Trypsin activity (a), TAP levels (b), and LDH leakage (c) were measured. Values are mean ± SEM obtained from at least three independent experiments and are expressed as percent of maximal response to cerulein stimulation in freshly prepared pancreas fragments. AP < 0.05 when cerulein-treated freshly prepared fragment values are compared with the basal values. BP < 0.05 when antisense oligonucleotide-incubated cerulein-stimulated fragments are compared with cerulein-stimulated 12-hour control fragments. (d) Quercetin treatment which blocks HSP70 induction restored caerulein induced trypsinogen activation. (Reproduced with permission from J Clin Invest) (4)
HSP27 is another inducible stress protein, whose protective role has been studied in acute pancreatitis. Kubisch et al reported that HSP27 overexpression in the pancreas also protects against caerulein-induced pancreatitis (18). In this study, huHSP27 was overexpressed under the control of CMV promoter. Of all the tissues examined, high expression of huHSP27 was found in pancreas and stomach. The protection was mediated by phosphorylated form of HSP27 because when non-phosphorylatable huHSP27 mutant form was used, the effect was not observed. HSP27 differs from other HSPs in that it modulates actin dynamics following HSP27 phosphorylation. Stabilization of microfilaments is thought to be responsible for an increased survival of cells recovering from cell stress.
While, there is overwhelming evidence to indicate that overexpression of HSP70, -60 or -27 has a protective effect on pancreatitis and that this effect is observed irrespective of method used for inducing overexpression or the experimental model used for induction of pancreatitis, however in an interesting study by Lunova et al (23), acinar cell specific overexpression of HSP70 did not protect against severity of pancreatitis but did accelerate the recovery from pancreatitis.

Figure 3. Effects of prior thermal stress and oligonucleotide treatment on morphologic changes induced by supramaximal cerulein in rat pancreas. Representative light micrographs of the H&E-stained pancreas sections from (A) control, (B) CER-, (C) H+CER-, (D) H+AS+CER-, (E) H+S+CER-, and (F) AS+CER-treated rats are shown. Note increased vacuolization/necrosis and leukocyte infiltration in CER and H+AS+CER groups. Both H+CER- and H+S+CER-treated rats show marked protection, whereas (F) animals given AS-HSP70 before administration of cerulein show more pronounced acinar cell vacuolization/necrosis. (Reproduced with permission from Gastroenterology) (5)
This somewhat contradictory finding could be explained by the observation that HSP70 ATPase activity is slow but in presence of its co-chaperone HSP40, it is accelerated many folds and this protein also help in binding of HSP70 to polypeptide. Efficient cleavage of ATP and effective binding to unfolded polypeptides determine the effectiveness of HSP70. In targeted overexpression of HSP70, availability of co-chaperone may become a limitation, which may not be the case in induction of stress proteins by other methods.
4. Mechanism of HSP overexpression mediated protection
The mechanism by which HSP overexpression provides protection against injury in pancreatitis has been investigated in studies where either HSP70, HSP60 or HSP27 was overexpressed. In the studies, where HSP70 was overexpressed, either using heat stress or by pharmacological means, it significantly inhibited activation of trypsinogen to trypsin during initiation of pancreatitis (Figure 2 & 4). Activation of trypsin is observed early on and is considered to be the key pathological event (4, 5) in acute pancreatitis. Inhibition of trypsin activation by different means have been shown to be protective (11, 33, 39)while disrupting the protective mechanisms by deletion of SPINK1 (17, 26) or Chymotrypsin C (36)accentuate trypsin activation and pancreatitis. The mechanism by which HSP70 prevents trypsin activation has also been investigated. Studies suggest that HSP70 overexpression interferes with co-localization of zymogens and lysosomes, which is prerequisite for intracellular activation of trypsinogen to trypsin. In non-pancreatic cells HSP70 has been suggested to influence calcium homeostasis (16). Given the requirement of Ca2+ for co-localization of zymogen and lysosomal contents and evidence of HSP70 perturbing Ca2+ signal, HSP70 mediated inhibition of co-localization could be through attenuation of pathological Ca2+ signaling. Another key event in the pathogenesis of acute pancreatitis is activation of nuclear factor kappa B (NFkB) which is independent of trypsin activation and contributes to development of pancreatitis and associated systemic injury. It has been shown that overexpression of HSP70 also provides protection against activation of NFkB (3) (Figure 5).

Figure 4. Effect of sodium arsenite pretreatment on cerulein-induced co-localization and trypsinogen activation. A: Subcellular distribution of cathepsin B. Rats were given a single intraperitoneal injection (10 mg/kg) of sodium arsenite (ARS). After 14 h, pancreatitis was induced in both treated and untreated rats by a single injection of cerulein (CER, 20 µg/kg); rats were sacrificed 2.5 h later. Untreated and ARS alone-treated rats served as controls (CON). Cathepsin B activity in subcellular fractions was measured as described in the text. Values are mean ± SEM for at least three independent experiments with at least three or four rats per group in an experiment. *P < 0.05 when ARS + CER rats were compared with CER rats alone. (B) and (C) trypsinogen activation. as measured as trypsin (B) and TAP levels (C). Values are mean ± SEM for three independent experiments. *P < 0.05 when ARS + CER rats were compared with the CER rats. (Reproduced with permission from J Cell Physiol) (3)
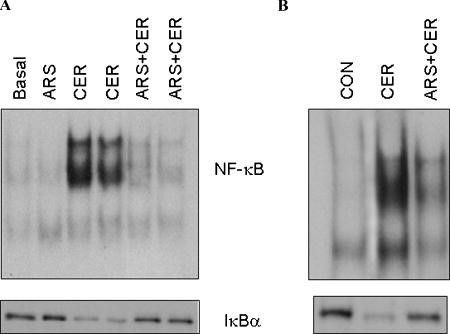
Figure 5. Pre-induction of heat shock protein 70 expression using sodium arsenite inhibited secretagogue‐induced NF‐κB activation in pancreas. Upper panel in A and B is EMSA of nuclear fraction, while lower panels depict IkBa degradation in cytosolic fraction. (Reproduced with permission from J Cell Physiol) (3)
Besides the mechanisms described above, other mechanisms of HSP mediated protection in acute pancreatitis have been proposed. All three major components of cytoskeleton namely microfilaments, intermediate filaments, and microtubules are present in pancreatic acinar cells (37). Stimulation of acinar cells with supramaximal concentration of caerulein or induction of acute pancreatitis in vivo results in loss of filamentous actin. Arginine administration has also been reported to cause changes in the actin cytoskeleton, including reduced actin staining under the luminal membrane and increased cytoplasmic staining (37). These changes in cytoskeleton are considered early changes related to acinar cell injury in pancreatitis. It has been suggested that HSP27 overexpression provide protection through regulation of actin (37). This stabilization of microfilaments is considered to be important for an increased survival of cells recovering from cell stress. The protective effects of HSP27 overexpression correlates with not only overexpression but also with phosphorylation. Overexpression of HSP70 have also been shown to protect cytoskeleton. The stress conditions, which induce HSP27 are also known to cause increase in HSP70 (37). The observed protection of HSP27 to cytoskeleton may also be enhanced in presence of HSP70 overexpression.
It has also been reported by Park et al (28) that HSP27 associates with IkB kinase complex and this interaction is stimulated by treatment with tumor necrosis factor alpha. There are contradictory reports about enhanced association of phosphorylated HSP27 with IkB kinase complex and resulting decreased IkB kinase activity because no increased activity was observed in transgenic mice with overexpression of HSP27 (18).
Overexpression of HSP27 also didn’t affect intracellular calcium release in response to stimulation with caerulein. Thus the protective effect of HSP27 overexpression during acute pancreatitis is mainly related to cytoskeleton stabilization. Subcellular redistribution of cathepsin B from lysosome-enriched fraction to the zymogen granule-enriched fraction is decreased in the group subjected to water-immersion which indicate HSP60 mediated protection might be because of interference with co-localization and resulting decrease in trypsin activation.
5. Summary
There is strong experimental evidence to support, that HSPs are overexpressed during development of pancreatitis. Increased expression of HSPs is a protective response, and is observed in multiple models of experimental pancreatitis. Pre-induction of HSPs by different means protects from subsequent pancreatitis induced injury. Mechanism of protection is not fully understood but prevention of intra-acinar trypsinogen activation, cytoskeleton stabilization and inhibition of NFkB signaling appears to be the main contributors. Better and safe pharmacological approaches for induction of HSPs could help in reducing severity of pancreatitis and related complications.
6. References
- Balog A, Gyulai Z, Boros LG, Farkas G, Takacs T, Lonovics J, et al. Polymorphism of the TNF-alpha, HSP70-2, and CD14 genes increases susceptibility to severe acute pancreatitis. Pancreas 30(2): e46-50, 2005. PMID: 15714129.
- Benndorf R, Hayess K, Ryazantsev S, Wieske M, Behlke J and Lutsch G. Phosphorylation and supramolecular organization of murine small heat shock protein HSP25 abolish its actin polymerization-inhibiting activity. J Biol Chem 269(32): 20780-20784, 1994. PMID: 8051180.
- Bhagat L, Singh VP, Dawra RK and Saluja AK. Sodium arsenite induces heat shock protein 70 expression and protects against secretagogue-induced trypsinogen and NF-kappaB activation. J Cell Physiol 215(1): 37-46, 2008. PMID: 17941083.
- Bhagat L, Singh VP, Hietaranta AJ, Agrawal S, Steer ML and Saluja AK. Heat shock protein 70 prevents secretagogue-induced cell injury in the pancreas by preventing intracellular trypsinogen activation. J Clin Invest 106(1): 81-89, 2000. PMID: 10880051.
- Bhagat L, Singh VP, Song AM, van Acker GJ, Agrawal S, Steer ML, et al. Thermal stress-induced HSP70 mediates protection against intrapancreatic trypsinogen activation and acute pancreatitis in rats. Gastroenterology 122(1): 156-165, 2002. PMID: 11781290.
- Cheng MY, Hartl FU and Horwich AL. The mitochondrial chaperonin hsp60 is required for its own assembly. Nature 348(6300): 455-458, 1990. PMID: 1978929.
- Choi HI, Lee SP, Kim KS, Hwang CY, Lee YR, Chae SK, et al. Redox-regulated cochaperone activity of the human DnaJ homolog Hdj2. Free Radic Biol Med 40(4): 651-659, 2006. PMID: 16458196.
- Deng M, Chen PC, Xie S, Zhao J, Gong L, Liu J, et al. The small heat shock protein alphaA-crystallin is expressed in pancreas and acts as a negative regulator of carcinogenesis. Biochim Biophys Acta 1802(7-8): 621-631, 2010. PMID: 20434541.
- Friant S, Meier KD and Riezman H. Increased ubiquitin-dependent degradation can replace the essential requirement for heat shock protein induction. EMBO J 22(15): 3783-3791, 2003. PMID: 12881413.
- Frossard JL, Saluja AK, Mach N, Lee HS, Bhagat L, Hadenque A, et al. In vivo evidence for the role of GM-CSF as a mediator in acute pancreatitis-associated lung injury. Am J Physiol Lung Cell Mol Physiol 283(3): L541-548, 2002. PMID: 12169573.
- Halangk W, Lerch MM, Brandt-Nedelev B, Roth W, Ruthenbuerger M, Reinheckel T, et al. Role of cathepsin B in intracellular trypsinogen activation and the onset of acute pancreatitis. J Clin Invest 106(6): 773-781, 2000. PMID: 10995788.
- Halangk W, Matthias R, Nedelev B, Schild L, Meyer F, Schulz HU, et al. [Modification of energy supply by pancreatic mitochondria in acute experimental pancreatitis]. Zentralbl Chir 122(4): 305-308, 1997. PMID: 9221643.
- Hohfeld J and Hartl FU. Role of the chaperonin cofactor Hsp10 in protein folding and sorting in yeast mitochondria. J Cell Biol 126(2): 305-315, 1994. PMID: 7913473.
- Jackson SE. Hsp90: structure and function. Top Curr Chem 328: 155-240, 2013. PMID: 22955504.
- Jia H, Halilou AI, Hu L, Cai W, Liu J and Huang B. Heat shock protein 10 (Hsp10) in immune-related diseases: one coin, two sides. Int J Biochem Mol Biol 2(1): 47-57, 2011. PMID: 21969171.
- Kiang JG, Ding XZ and McClain DE. Overexpression of HSP-70 attenuates increases in [Ca2+]i and protects human epidermoid A-431 cells after chemical hypoxia. Toxicol Appl Pharmacol 149(2): 185-194, 1998. PMID: 9571987.
- Koziel D, Gluszek S, Kowalik A, Chlopek M and Pieciak L. Genetic mutations in SPINK1, CFTR, CTRC genes in acute pancreatitis. BMC Gastroenterol 15: 70, 2015. PMID: 26100556.
- Kubisch C, Dimagno MJ, Tietz AB, Welsh MJ, Ernst SA, Brandt-Nedelev B, et al. Overexpression of heat shock protein Hsp27 protects against cerulein-induced pancreatitis. Gastroenterology 127(1): 275-286, 2004. PMID: 15236192.
- Lee HS, Bhagat L, Frossard JL, Hietaranta A, Singh VP, Steer ML, et al. Water immersion stress induces heat shock protein 60 expression and protects against pancreatitis in rats. Gastroenterology 119(1): 220-229, 2000. PMID: 10889172.
- Li J, Soroka J and Buchner J. The Hsp90 chaperone machinery: conformational dynamics and regulation by co-chaperones. Biochim Biophys Acta 1823(3): 624-635, 2012. PMID: 21951723.
- Lindquist S. The heat-shock response. Annu Rev Biochem 55: 1151-1191, 1986. PMID: 2427013.
- Lindquist S and Craig EA. The heat-shock proteins. Annu Rev Genet 22: 631-677, 1988. PMID: 2853609.
- Lunova M, Zizer E, Kucukoglu O, Schwarz C, Dillmann WH, Wagner M, et al. Hsp72 overexpression accelerates the recovery from caerulein-induced pancreatitis. PLoS One 7(7): e39972, 2012. PMID: 22792201.
- Mayer MP and Bukau B. Hsp70 chaperones: cellular functions and molecular mechanism. Cell Mol Life Sci 62(6): 670-684, 2005. PMID: 15770419.
- Miron T, Vancompernolle K, Vandekerckhove J, Wilchek M and Geiger B. A 25-kD inhibitor of actin polymerization is a low molecular mass heat shock protein. J Cell Biol 114(2): 255-261, 1991. PMID: 2071672.
- Nathan JD, Romac J, Peng RY, Peyton M, Macdonald RJ and Liddle RA. Transgenic expression of pancreatic secretory trypsin inhibitor-I ameliorates secretagogue-induced pancreatitis in mice. Gastroenterology 128(3): 717-727, 2005. PMID: 15765407.
- Parag HA, Raboy B and Kulka RG. Effect of heat shock on protein degradation in mammalian cells: involvement of the ubiquitin system. EMBO J 6(1): 55-61, 1987. PMID: 3034579.
- Park KJ, Gaynor RB and Kwak YT. Heat shock protein 27 association with the I kappa B kinase complex regulates tumor necrosis factor alpha-induced NF-kappa B activation. J Biol Chem 278(37): 35272-35278, 2003. PMID: 12829720.
- Plusczyk T, Westermann S, Rathgeb D and Feifel G. Acute pancreatitis in rats: effects of sodium taurocholate, CCK-8, and Sec on pancreatic microcirculation. Am J Physiol Gastrointest Liver Physiol 272(2 Pt 1): G310-320, 1997. PMID: 9124355.
- Qiu XB, Shao YM, Miao S and Wang L. The diversity of the DnaJ/Hsp40 family, the crucial partners for Hsp70 chaperones. Cell Mol Life Sci 63(22): 2560-2570, 2006. PMID: 16952052.
- Ritossa F. A new puffing pattern induced by temperature shock and DNP in drosophila. Experientia 18(12): 571-573, 1962.
- Rogalla T, Ehrnsperger M, Preville X, Kotlyarov A, Lutsch G, Ducasse C, et al. Regulation of Hsp27 oligomerization, chaperone function, and protective activity against oxidative stress/tumor necrosis factor alpha by phosphorylation. J Biol Chem 274(27): 18947-18956, 1999. PMID: 10383393.
- Saluja AK, Donovan EA, Yamanaka K, Yamaguchi Y, Hofbauer B and Steer ML. Cerulein-induced in vitro activation of trypsinogen in rat pancreatic acini is mediated by cathepsin B. Gastroenterology 113(1): 304-310, 1997. PMID: 9207291.
- Schirmer EC, Glover JR, Singer MA and Lindquist S. HSP100/Clp proteins: a common mechanism explains diverse functions. Trends Biochem Sci 21(8): 289-296, 1996. PMID: 8772382.
- Stetler RA, Gao Y, Zhang L, Weng Z, Zhang F, Hu X, et al. Phosphorylation of HSP27 by protein kinase D is essential for mediating neuroprotection against ischemic neuronal injury. J Neurosci 32(8): 2667-2682, 2012. PMID: 22357851.
- Szabo A, Ludwig M, Hegyi E, Szepeova R, Witt H and Sahin-Toth M. Mesotrypsin Signature Mutation in a Chymotrypsin C (CTRC) Variant Associated with Chronic Pancreatitis. J Biol Chem 290(28): 17282-17292, 2015. PMID: 26013824.
- Tashiro M, Schafer C, Yao H, Ernst SA and Williams JA. Arginine induced acute pancreatitis alters the actin cytoskeleton and increases heat shock protein expression in rat pancreatic acinar cells. Gut 49(2): 241-250, 2001. PMID: 11454802.
- Tissieres A, Mitchell HK and Tracy UM. Protein synthesis in salivary glands of Drosophila melanogaster: relation to chromosome puffs. J Mol Biol 84(3): 389-398, 1974. PMID: 4219221.
- Van Acker GJ, Saluja AK, Bhagat L, Singh VP, Song AM and Steer ML. Cathepsin B inhibition prevents trypsinogen activation and reduces pancreatitis severity. Am J Physiol Gastrointest Liver Physiol 283(3): G794-800, 2002. PMID: 12181196.
- Voronina SG, Barrow SL, Simpson AW, Gerasimenko OV, da Silva Xavier G, Rutter GA, et al. Dynamic changes in cytosolic and mitochondrial ATP levels in pancreatic acinar cells. Gastroenterology 138(5): 1976-1987, 2010. PMID: 20102715.
- Wagner AC, Weber H, Jonas L, Nizze H, Strowski M, Fiedler F, et al. Hyperthermia induces heat shock protein expression and protection against cerulein-induced pancreatitis in rats. Gastroenterology 111(5): 1333-1342, 1996. PMID: 8898648.
- Weber CK, Gress T, Muller-Pillasch F, Lerch MM, Weidenbach H and Adler G. Supramaximal secretagogue stimulation enhances heat shock protein expression in the rat pancreas. Pancreas 10(4): 360-367, 1995. PMID: 7792292.
- Welch WJ. Mammalian stress response: cell physiology, structure/function of stress proteins, and implications for medicine and disease. Physiol Rev 72(4): 1063-1081, 1992. PMID: 1438579.