
Entry Version:
Citation:
Pancreapedia: Exocrine Pancreas Knowledge Base, DOI: 10.3998/panc.2021.01
| Attachment | Size |
|---|---|
| 4.79 MB | |
| 3 MB |
I. Introduction
The mandate for this chapter is to review the anatomy and histology of the pancreas. The pancreas (meaning all flesh) lies in the upper abdomen behind the stomach. The pancreas is part of the gastrointestinal system that makes and secretes digestive enzymes into the intestine, and also an endocrine organ that makes and secretes hormones into the blood to control energy metabolism and storage throughout the body.
It is worthwhile to mention a few definitions for key terms as used in the context of the pancreas:
Exocrine pancreas, the portion of the pancreas that makes and secretes digestive enzymes into the duodenum. This includes acinar and duct cells with associated connective tissue, vessels, and nerves. The exocrine components comprise more than 95% of the pancreatic mass.
Endocrine pancreas, the portions of the pancreas (the islets) that make and secrete insulin, glucagon, somatostatin and pancreatic polypeptide into the blood. Islets comprise 1-2% of the pancreatic mass.
Since we are dealing with a three dimensional solid structure, the aphorism that “a picture is worth a thousand words” seems to pertain (1). Accordingly, this chapter will largely consist of images with extended legends. The images range from classic work of skilled medical artists to original drawings and photomicrographs from leaders in the study of pancreatic anatomy and structure. Text is interspersed as appropriate. Additional useful images are available online at other websites. We provide a list of some of these sites at the end with the references.
II. Gross Anatomy
Figures 1-13 depict the gross anatomy of the pancreas and its relationship to surrounding organs in adults. It is customary to refer to various portions of the pancreas as head, body, and tail. The head lies near the duodenum and the tail extends to the hilum of the spleen.
When the terms anterior, posterior, front and back are used, they pertain to relationships in the human, standing erect. Superior and inferior are used in the same context so that they mean toward the head and toward the feet, respectively. These usages obviously do not pertain in quadruped animals where dorsal, ventral, cephalad, and caudad are more useful terms.
Use of the terms left and right can be problematic. For example, the spleen is located in the upper portion of the abdomen on the left side of the body. When the abdomen is pictured from the front, this places the spleen on the viewer’s right hand side. We will adopt the convention that right and left (unqualified) will be used in the first sense in the legends for gross anatomy (indicating the subject’s right and left side).
Figure 1. The gross anatomy of the human pancreas can vary. Figures 1A and 1B are two normal human pancreases from autopsies of adults. Both pancreases have been dissected to remove fat and adjacent organs. The two photos illustrate that there is considerable individual variation in the shape of the pancreas.
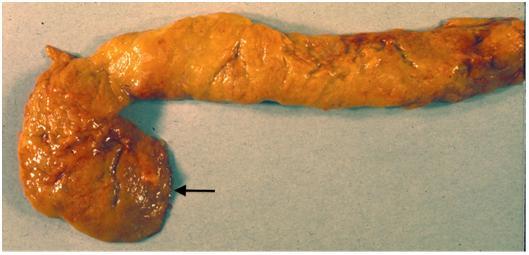
(A) This pancreas has a conspicuous uncinate lobe that curves down and to the left (arrow). This is an unusual configuration since the uncinate process usually fuses more completely with the dorsal pancreas adding mass to the head of the pancreas as seen in Figures. 1B and 2.

(B) In this pancreas the uncinate portion is fused to the remainder of the head. A probe (image left) has been put into the main pancreatic duct, and a second probe (vertical) is in the portal-superior mesenteric vein behind the pancreas. The diagonal groove in the tip of the tail (image right) marks the course of a branch of the splenic artery or vein.
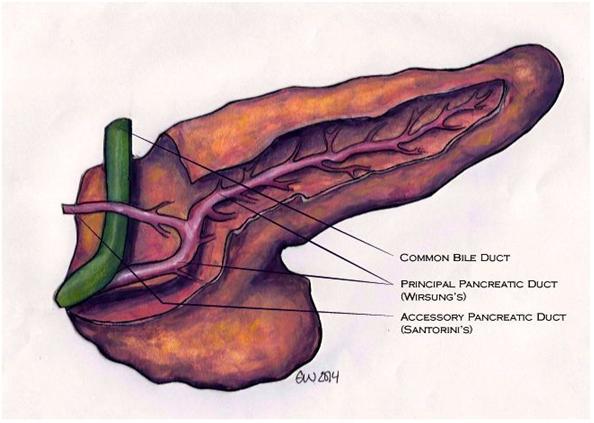
Figure 2. Normal pancreas dissected to reveal the duct system. The pancreas is viewed from the front and a portion of the parenchyma has been dissected away to reveal (1) the main (principal) pancreatic duct (Wirsung’s duct) with multiple branches, (2) the accessory duct (Santorini’s duct), and (3) the distal common bile duct. Although the regions are not labeled we see the head of the pancreas at image left, and tail of the pancreas, image right. This drawing depicts a configuration that is intermediate to those shown in Figure 1 in regard to the degree of fusion of the uncinate process with the dorsal pancreas. (Drawing by Emily Weber for Pancreapedia).
When we are designating location within an image, we will use “image right” and “image left” to denote relationships within the image.
Artwork in Figures 3, 7-8, and 11-13 is by Jennifer Parsons Brumbaugh. These drawings were originally published in the AFIP Fascicle on pancreatic neoplasms and are used with permission of the publisher (6). Chapter 1 of the Fascicle is recommended as a source for additional detail regarding pancreatic anatomy and histology, and for discussion of the genetic control of pancreatic development.
The tail of the pancreas and spleen are in the left upper quadrant of the abdomen and the head of the pancreas is in the right upper quadrant just to the right of the midline. If you place your right hand over your upper abdomen with fingers extending to the left over the lower portion of your rib cage and the tip of your thumb extended up over the lower portion of the sternum, then your pancreas lies behind your hand in the back (retroperitoneal) portion of the abdomen. This may be visualized by reference to the small image in the upper image right corner in Figure 4. The pancreas is about the size of the half of your hand that includes the index and third fingers excluding the thumb. The pancreas weighs about 100 grams and is 14-23 cm long (6).
Figure 3. Anatomic relationships of the pancreas with surrounding organs and structures Several key relationships should be noted. Their recognition may be facilitated by also referring to Figures 4 and 5.
• The head of the pancreas lies in the loop of the duodenum as it exits the stomach.
• The tail of the pancreas lies near the hilum of the spleen.
• The body of the pancreas lies posterior to the distal portion of the stomach between the tail and the neck and is unlabeled in this drawing.
• The portion of the pancreas that lies anterior to the aorta is somewhat thinner than the adjacent portions of the head and body of the pancreas. This region is sometimes designated as the neck of the pancreas and marks the junction of the head and body.
• The close proximity of the neck of the pancreas to major blood vessels posteriorly including the superior mesenteric artery, superior mesenteric-portal vein, inferior vena cava, and aorta limits the option for a wide surgical margin when pancreatectomy (surgical removal of the pancreas) is done.
• The common bile duct passes through the head of the pancreas to join the main duct of the pancreas near the duodenum as shown in Figure 2. The portion nearest the liver lies in a groove on the dorsal aspect of the head (see Figure 7B).
• The minor papilla where the accessory pancreatic duct drains into the duodenum and the major papilla (ampulla of Vater) where the main pancreatic duct enters the duodenum are depicted, image left. Image by Jennifer Parsons Brumbaugh; used with permission of the publisher (6)).

Figure 4. Cross section of the upper abdomen at the level of the pancreas. Note that the plane of the transection is angled upward on the left as indicated in the drawing upper image right. The major organs except the liver (image left) and kidneys are labeled. Splenic flexure (image right) refers to the colon. Used with permission. Copyright, American Gastroenterological Association.
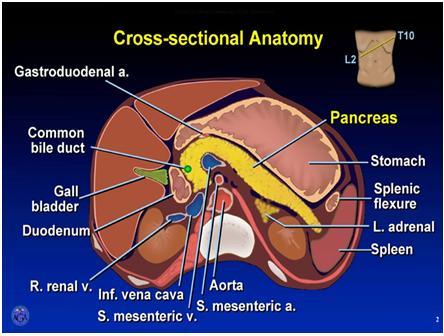
Figure 5. CT scan of the upper abdomen at the level of the pancreas. This annotated CT scan is oriented with the abdominal wall at the top and the spine and muscles of the back at the bottom viewing the cross section from below. Thus, the spleen is at the extreme image right and the liver is image left inside the ribs that appear as white ovals in the abdominal wall. Kidneys lie lateral to the spinal column with the tail of the pancreas nearly touching the left kidney. (Used with permission. Copyright, American Gastroenterological Association).

Figure 6. Mouse pancreas. The pancreas of an adult mouse is shown surrounded by the stomach (top), the duodenum and proximal jejunum (image left and bottom), and the spleen (image right). The duodenum wraps around the head of the pancreas (as demarcated by the line). Rodent pancreas is soft and diffuse compared with the human pancreas. (Photo provided by Catherine Carriere).

Figure 7. The arterial blood supply of the pancreas. The upper panel (A) is visualized from the front, and the lower panel (B) is seen from the back. The celiac trunk and the superior mesenteric artery both arise from the abdominal aorta. Both have multiple branches that supply several organs including the pancreas. The anastomosis of their branches around the pancreas provides collateral circulation that generally assures a secure arterial supply to the pancreas. Most of the arteries are accompanied by veins (not shown) that drain into the portal and splenic veins as they pass behind the pancreas as shown in B. The superior mesenteric vein becomes the portal vein when it joins the splenic vein. (Image by Jennifer Parsons Brumbaugh used with permission of the publisher (6)).
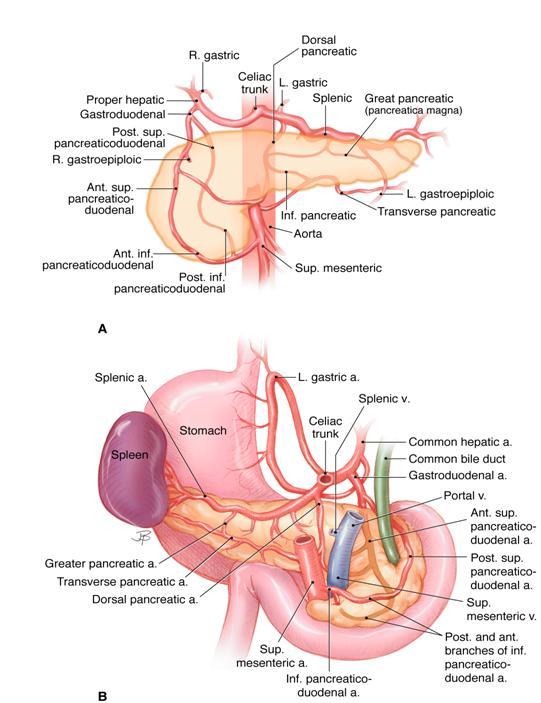
Figure 8. Lymph nodes draining the pancreas. This figure indicates the typical location of lymph nodes surrounding the pancreas. There is considerable individual variation in the location of lymph nodes and an image like this is idealized. Both A and B are anterior views. B includes some nodes that lie posterior to the pancreas. (Image by Jennifer Parsons Brumbaugh used with permission of the publisher (6)).

Figure 9. Pancreas-associated lymph nodes are assigned numerical codes (lymph node station numbers) that correspond to their anatomic location. This classification is used to denote the location of metastatic spread of pancreatic neoplasms or for other detailed studies. These station numbers are seldom used in Western publications and the image is provided primarily for reference. (Figure used with permission of the Japan Pancreas Association and the Kanehara publishers).
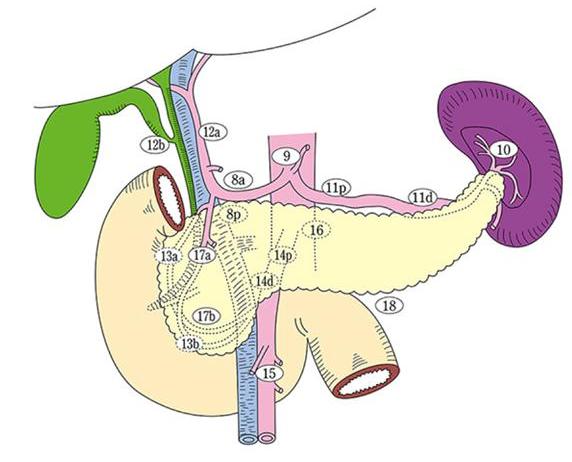
Figure 10. Nerves (yellow) serving the pancreas. The cross sectional image (A) emphasizes the location of the celiac ganglia of the autonomic system lateral to the aorta while (B) emphasizes the rich nerve plexus that connects these ganglia to the pancreas. SMA (superior mesenteric artery). PL (plexus). (Figure used with permission of the Japan Pancreas Association and the Kanehara publishers).

There is no anatomic landmark for the division between the body and tail of the pancreas although the left border of the aorta is sometimes used to mark the junction (2, 6). Hellman defined the tail as the one fourth of the pancreas from the tip of the tail to the head, whereas Wittingen defined the junction between the body and tail as the point where the gland sharply narrowed (4, 16). It would be difficult to define this point in the pancreases shown in Figure 1.
III. Embryology and Development
The pancreatobiliary anlagen appear at gestation week 5 in the human; fusion of the dorsal and ventral anlagen occurs during week 7 (10). Full development of acinar tissue extends into the postnatal period. In mice, pancreatic development begins at embryonic day 8.5 (e8.5) and is largely complete by day e14.5 (7).
Figure 11. (A) The embryonic development of the pancreas and biliary system in the human. (B) The rotation of the ventral anlage to fuse with the dorsal anlage and fusion of the duct systems such that the main pancreatic duct is formed from portions of the ducts of both dorsal and ventral anlagen.
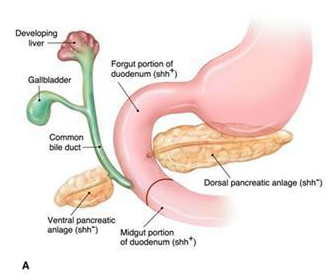
(A) The pancreas develops from two outgrowths of the foregut distal to the stomach. The ventral diverticulum gives rise to the common bile duct, gallbladder, liver and the ventral pancreatic anlage that becomes a portion of the head of the pancreas with its duct system including the uncinate portion of the pancreas. The dorsal pancreatic anlage gives rise to a portion of the head, the body, and tail of the pancreas including a major duct that is continuous through the three regions.
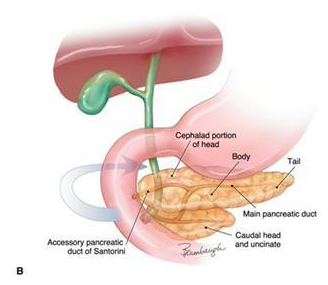
(B) The caudal portion of the head of the pancreas (uncinate) and the major papilla (ampulla of Vater) are derived from the ventral anlage. The minor papilla that drains the duct of Santorini is derived from the dorsal anlage. (Image by Jennifer Parsons Brumbaugh; used with permission of the publisher (6)).
Figure 12. Anatomic variations in the pancreatic and common bile duct systems. The anatomic variations depicted provide additional examples of individual differences in pancreatic anatomy seen in adults. These will be most easily understood by comparing Figures 11 and 12. It becomes apparent that the duct of Santorini is derived from the dorsal anlage, whereas the duct of Wirsung (the main duct of the pancreas) is derived from the fusion of duct systems of both dorsal and ventral anlagen and drains into the duodenum at the ampulla of Vater as depicted in A and B. The connection of the duct of Santorini to the duodenum may regress as depicted in A or persist as in B, C, and D. The duct systems of the two anlagen may fail to fuse as depicted in C giving rise to “pancreas divisum”. Rarely the duct systems may fuse but lose their connection to the ampulla as depicted in D. Pancreatic secretions then reach the duodenum through the duct of Santorini and the minor papilla. (Image by Jennifer Parsons Brumbaugh; used with permission of the publisher (6)).
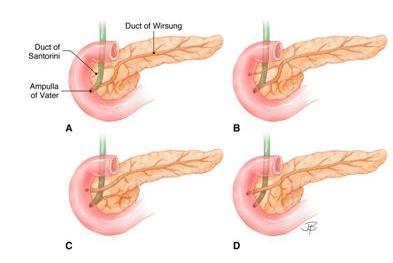
Eponymic names identify the anatomist, embryologist or physician who is credited with first describing a structure. You may conclude that Wirsung, Santorini, and Vater were such scientists.
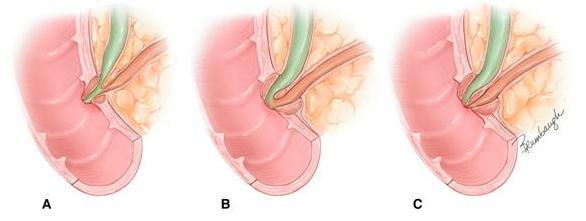
Figure 13. Anatomic variations in the union of the common bile duct and the main pancreatic duct at the major papilla (ampulla of Vater). “Common channel” refers to the fused portion of the bile and pancreatic ducts proximal to entry into the duodenum. The common channel may be long as depicted in A or short as in B. Less often, there is no common channel because the ducts open separately into the duodenum as depicted in C. The common channel has received much attention because stones in the biliary tract (gallstones) may lodge in the common channel causing obstruction of both pancreatic and biliary duct systems. Such an obstruction is frequently regarded as the cause of acute pancreatitis. (Image by Jennifer Parsons Brumbaugh; used with permission of the publisher (6)).
IV. Histology and Ultrastructure
Figures 14-29 depict the histology of the exocrine pancreas at the light and electron microscopic levels. Most histologic images are from human tissue. Exceptions are usually noted in the legend. In Hematoxylin and Eosin (H&E) stained sections, nucleic acids (DNA and RNA) stain blue; most proteins and carbohydrates stain pink to red; fat is extracted by organic solvents used in tissue processing leaving unstained spaces. Sections for light microscopy are most often made from formalin-fixed paraffin-embedded tissue and the sections are usually 4 or 5 micrometers (μm) thick. Thinner (1 μm) sections of plastic embedded tissues (prepared for electron microscopy) may also be used for light microscopy and a few such sections are also illustrated. For additional ultrastructural detail the reader is referred to the chapter by Kern (8).
Figure 14. Fetal pancreas (H&E). This tissue section illustrates developing exocrine tissue in the center (arrows) surrounded by primitive mesenchymal and hematopoietic cells at an estimated gestational age of 5 weeks. The acinar tissue is composed of a network of interconnecting tubules. (Micrograph contributed by Dale E. Bockman).
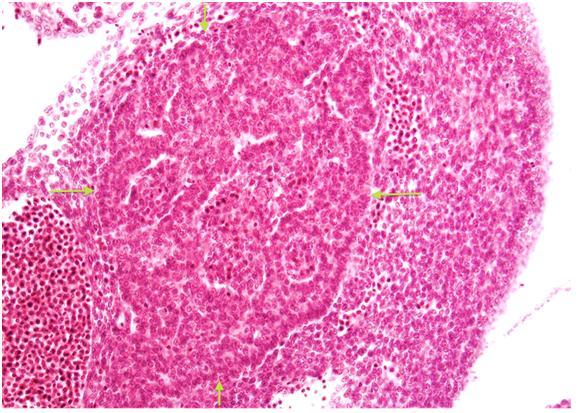
Figure 15. The exocrine pancreas is a complex tubular network. The point of this drawing is that pancreatic acini are not arranged in clusters like grapes at the ends of a branching duct system but rather as an anastomosing tubular network that at some termini form classic acini. Centroacinar cells are typically located at the junction of an acinus or acinar tubule with a small ductule, but they may be interspersed within an acinar tubule. In this drawing many acinar cells have been replaced by duct cells. This process, called acinar to ductal metaplasia (ADM), occurs in chronic pancreatitis (3). (Also see Figure 8 in (2)). (Image contributed by Dale E. Bockman).
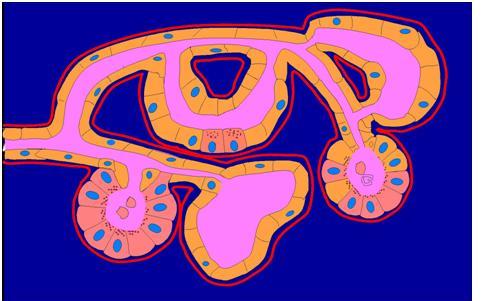
Figure 16. Acinar tissue, adult human pancreas (H&E). Acinar cells stain blue at their base because of the high content of RNA and the presence of nuclei. They are pink at their apex (lumenal aspect) where there is a high content of zymogen proteins (digestive enzymes). The nuclei of centroacinar cells are sometimes seen within an acinus (arrows).
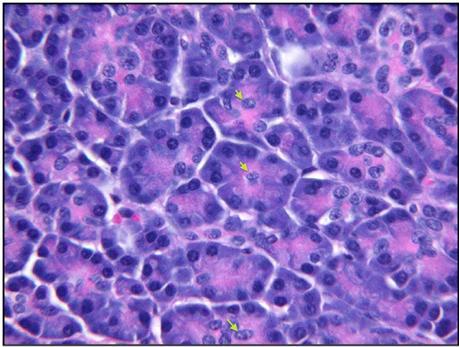
Figure 17. Pancreatic tissue with acinar, centroacinar and ductal cells (EM thick section). The acinar cells are larger than centroacinar cells and are easily identified because of the darkly stained zymogen granules (ZG). The basal portion (B) of the acinar cells lies next to the interstitial space that contains vessels (V), nerves and connective tissue. Nuclei (N) with nucleoli (n) are in the basal portion of the acinar cells. The golgi (G) lies at the junction of the basal and apical (A) portions of the cell. Centroacinar cells (CAC) have less rough endoplasmic reticulum and no secretory granules. Their cytoplasm is more lightly stained. A small ductule (D) extends from image right to below center. This is a 1 μm thick section of plastic embedded tissue prepared for electron microscopy that was stained with toluidine blue. (Micrograph contributed by James Jamieson).
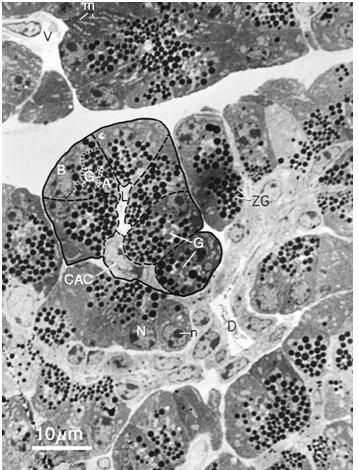
Figure 18. Pancreas with acinar and centroacinar cells with a small intralobular duct (Toluidine blue stain, 1 μm thick plastic embedded tissue). The presence of numerous round empty capillaries (arrows) in the interstitial spaces indicates that the pancreas was perfused with fixative. A small branching intralobular duct is evident at the top of the field. Blue zymogen granules are conspicuous in the acinar cells. (Micrograph contributed by James Jamieson).
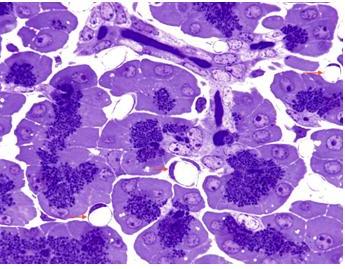
Figure 19. Acinar and centroacinar cells (low power electron micrograph). Zymogen granules, RER (rough endoplasmic reticulum), and nuclei are all identifiable in the acinar cells. In addition, several small dense inclusions of variable structure are present in the cytoplasm (lower red arrow). These are residual bodies derived from degradation of acinar cell organelles by lysosomal enzymes. The formation of such residual bodies is called autophagy, and large complex membrane-bound structures reflecting this process are called autophagic vacuoles. Such “cellular debris” is sometimes extruded into the interstitium as seen near the top of the field (upper red arrow). Residual bodies are also sometimes extruded into the acinar lumen providing a pathway for “garbage” disposal into the intestine. An acinar lumen is indicated by a small black arrow that lies between two centroacinar cells left of center. Figures 21 and 22 show acinar lumens at higher magnification. Zymogen granules vary in size from about 0.5-1.4 μm. (Micrograph contributed by James Jamieson).
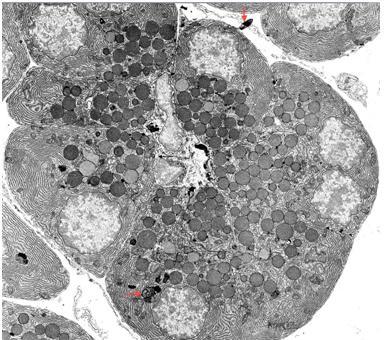
Figure 20. Rough endoplasmic reticulum (RER) shown by high magnification electron micrograph. The ribosomes adhere to the cytosolic surface of the membrane whereas the cisternal (luminal) side is devoid of ribosomes. Arrows in the cisterna (image left; red arrows) point toward the interior side of the endoplasmic reticulum. A few ribosomes appear to be free in the cytosol. (Micrograph by George Palade, contributed by James Jamieson).

Figure 21. Apical portions of acinar cells abutting two acinar lumens (electron micrograph). A portion of a centroacinar cell (CAC) forms part of the wall of the lower lumen (image right lower corner). The arrow in this lumen points to the CAC that has multiple mitochondria in the cytosol. Microvilli are evident protruding into the lumen from both CAC and acinar cells. A second smaller acinar lumen is near the image left upper corner. Zymogen granules are heavily stained so it is not possible to distinguish their membranes. RER is also evident in the acinar cells. (Micrograph contributed by James Jamieson).
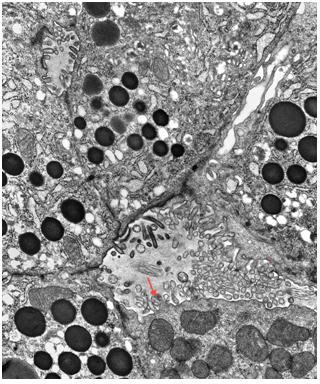
Figure 22. Apical domain of acinar cells is filled with zymogen granules (electron micrograph). The acinar cells abut a lumen near the center of the image. Microvilli protrude into the lumen. The section is lightly stained allowing visualization of the membrane of the zymogen granules. Zymogen granules are typically about 1 μm in diameter. Zonula occludens (tight junctions) are present near the acinar lumen (arrows). A mitochondrion is evident upper image left and a smaller one is located lower image left. (Micrograph contributed by James Jamieson).
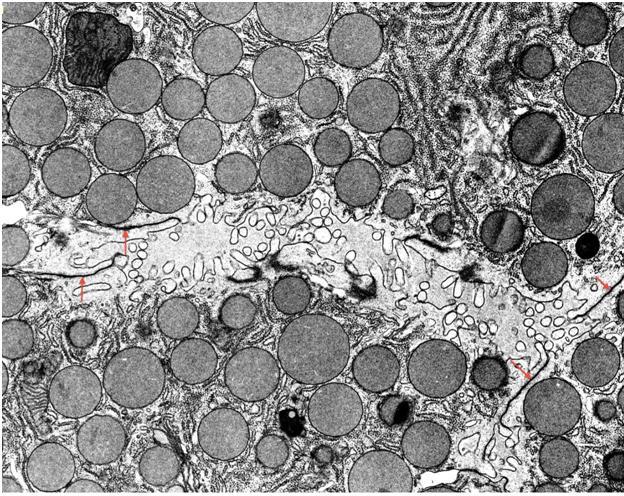
Figure 23. Key elements of the acinar cell protein synthetic pathway show a close physical relationship (Transmission EM). Many of the vesicles seen in the middle of the field are likely involved in the transport of newly synthesized proteins from the RER (image left) to the Golgi (right of center). Arrows mark budding of vesicles from the RER and indicate the direction of protein transport by the vesicles to the Golgi and thence to the formation of zymogen granules (image right). (Micrograph contributed by James Jamieson).
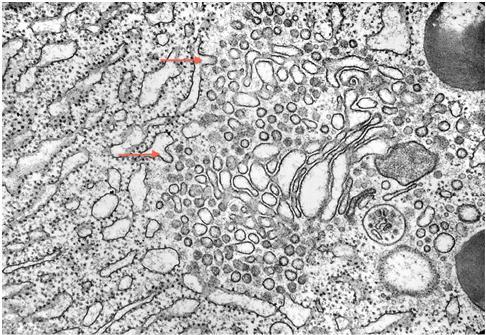
Figure 24. Steps of zymogen granule exocytosis at the apical membrane of the acinar cell are shown (Transmission EM). Right of center there is a zymogen granule with a hint of fusion of its membrane with the luminal cell membrane as an early step in secretion. To the left of this granule there is a “cup” in the cell surface that apparently marks the site of excretion of a zymogen granule after fusion of the membrane of zymogen granule with the luminal cell membrane. The secretory process has been described in detail (16). (Micrograph by George Palade, contributed by James Jamieson).

V. Duct System
The components of the duct system are the main pancreatic duct (duct of Wirsung), interlobular ducts that drain into the main duct throughout the pancreas as depicted in Figure 2, and intralobular ducts (sometimes called intercalated ductules) that link acinar tubules to the interlobular ducts. The intralobular ducts and ductules are ordinarily seen only at the level of light and electron microscopy. Enzymes from acinar cells are released into a bicarbonate-rich solution that is secreted by the centroacinar and ductal cells and flows from the acini and acinar tubules to the intralobular ducts, then into the interlobular ducts and main duct, and finally into the duodenum at the major or minor papillae. This duct system is illustrated in Figures 25-28.
The integrity of the duct system is of key importance in preventing entry of the exocrine enzymes into the interstitial space where they may be activated and cause tissue damage manifest as pancreatitis. The main and interlobular ducts have thick dense collagenous walls. The connective tissue component of the duct wall becomes progressively thinner as the ducts branch and become narrower. Intercellular tight junctions, also called zonula occludens, between duct cells, centroacinar cells and acinar cells play a major role in preventing leakage of the duct system. These have not been well illustrated although they can be seen in Figures 21 and 22 as dark, thickened zones in the adjacent cell membranes near the acinar or duct lumen. The chapter by Kern in The Pancreas provides excellent images and discussion of these tight junctions (8).
Figure 25. Main pancreatic duct, human. (H & E). The lumen is lined by a single layer of cuboidal duct cells. The thickness of the collagenous duct wall is impressive and is probably accentuated because the lumen is empty and collapsed.
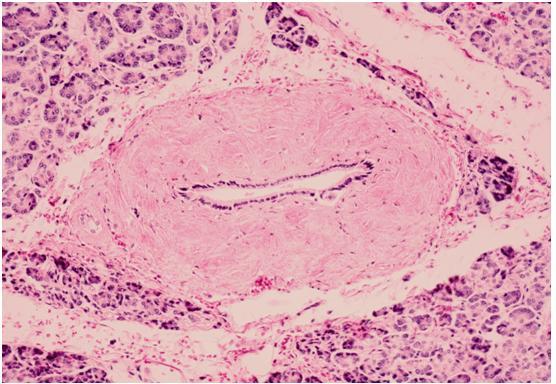
Figure 26. Interlobular duct, human (H & E). The lumen is lined by a single layer of duct cells. The collagenous wall is conspicuous but clearly thinner than that of the main duct. Near the center there is a smaller thin-walled intralobular duct joining the interlobular duct.

Figure 27. Intralobular ducts, human pancreas (H&E). An intralobular duct with a modest collagenous wall, image right, branches to give rise to an intralobular ductule that in turn branches, image left (arrow). The ductule is nearly devoid of collagen in its wall. The lumen of the small duct and ductule contains homogenous pink-staining protein-rich pancreatic juice. There is a small islet (small cells, pale cytoplasm) at the upper border, image left (asterisk).

Figure 28. Acinar tissue with an intralobular ductule in cross section, human pancreas (H & E). Note the single layer of cuboidal duct cells and the nearly complete absence of collagen in the wall of this ductule. Compare this with Figures 19 and 27, where intralobular ductules are shown in longitudinal section. The lumen of the ductule contains a pink granular proteinaceous precipitate from pancreatic juice. The clear spaces between the duct cells and the thin connective tissue wall of the ductule reflects artifactual separation of the cells from the basement membrane.
VI. Interstitial Tissue
Interstitial tissue surrounds lobules of acinar tissue, ducts and islets. The interstitium contains arteries, veins, capillaries, lymphatics, neural tissue, and stellate cells. Leukocytes may infiltrate the interstitium especially during pancreatitis or in reaction to neoplasms.
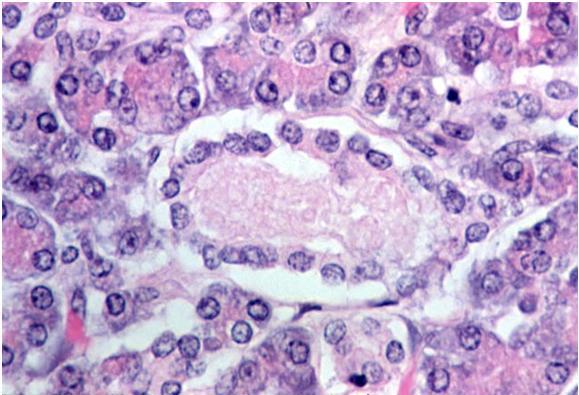
The pancreatic stellate cells (PSC) are specialized connective tissue cells with characteristic structure (Figure 29). They secrete multiple components of the extracellular matrix and are activated by a multitude of factors including inflammatory mediators, alcohol and its metabolites, endotoxins and cancer cell-derived factors. Activated PSCs function as fibroblasts, thus contributing to fibrosis associated with chronic pancreatitis and pancreatic ductal adenocarcinoma as discussed in detail by Mekapogu et al. (12).
Figure 29. Pancreatic stellate cell in the interstitial space surrounded by acinar cells. An extension of the cell’s cytoplasm extends downward between two acinar cells in the lower portion of the field. Dark inclusions in this extension are probably lipid droplets. (Micrograph contributed by the Pancreatic Research Group, UNSW, Australia; with special thanks to Dr. Murray Killingsworth).

VII. Endocrine Pancreas
Most islets (islets of Langerhans) that collectively comprise the endocrine pancreas are too small to be seen by gross examination, and thus they were not depicted in Figures 1-13. Islets vary greatly in size; ~70% are in the size range of 50-250 μm in diameter in humans with an average in the range of 100-150 μm (4). Smaller islets are dispersed throughout the acinar lobules and most larger islets lie along the main and interlobular ducts of the pancreas. Most islets are spherical or ellipsoid, but they can be irregular in shape--sometimes reflecting the pressure of an adjacent structure, often a duct, or limitation by a tissue plane. Several reports provide support for the presence of a higher population density of islets in the tail of the pancreas than in the head and body although others find no difference (5, 14, 17, 18). In adult humans the number of islets is calculated to be 500,000-1 million whereas there are far fewer in smaller animals (9, 11). Islets comprise 1-2% of the pancreas in adults of most mammalian species. In addition to the islets, isolated islet cells may be found dispersed in the acinar lobules or in association with ducts.
Photomicrographs of islets follow (Figures 30-38). Several of these have been immunostained using antibodies to specific islet peptide hormones to demonstrate various islet cell types including β-cells (insulin), α-cells (glucagon), δ-cells (somatostatin) and PP cells (pancreatic polypeptide). The PP-cells are commonly regarded as the fourth most prevalent endocrine cell type in the islets. Most PP-cells are in the portion of the pancreas derived from the ventral pancreatic anlage, i.e. the uncinate process that is reported to comprise about 10% of the pancreas (13, 15). In the portion of the pancreas derived from the dorsal pancreatic anlage, the majority of islet cells are β-cells (75-80%), followed by α-cells (about 15%), δ-cells (about 5%) and very few PP-cells. In the uncinate process there are few α-cells and many more PP-cells. Stefan et al. present data from study of non-diabetic human pancreases showing that the PP-cells comprise 54-94% of the volume of islets in the uncinate region, displacing most α-cells and some β-cells (15). These investigators provide data indicating that PP-cells are the second most prevalent endocrine cell type overall in the pancreas among their 13 nondiabetic subjects.
Figure 30. Human pancreas with three islets (H&E). This low power histologic section illustrates three islets in the background of the more abundant acinar tissue with a small duct in the upper image right corner. The large islet, image left, may be two adjacent islets with a small islet conforming to the lower border of a large round islet. Two small oval islets are located image right at 2 and 4 o’clock. The islet cells are smaller and have paler cytoplasm than the surrounding acinar cells.
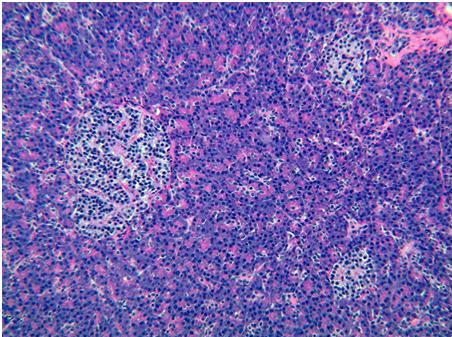
Figure 31. Human islet (H&E). This islet is elongate and nearly triangular in this cross section. A thin fibrous septum lies along its lower border. Although most islets are oval or round in cross section, islets vary greatly in shape as illustrated here.

Figure 32. Exocrine pancreas with an islet, mouse pancreas (H&E). The top of a large islet abuts an intralobular duct that is slightly left of center. (Photomicrograph by Catherine Carriere).

Figure 33. Islet cells store each hormone in distinct locations (Immunoperoxidase). Serial sections of an islet have been immunostained using antibodies to insulin (image left), glucagon (center) and somatostatin (image right). The presence of the hormones is indicated by the brown stain. The predominance of insulin secreting β-cells is obvious. In the center and image right photos, the location of α-cells and δ-cells is primarily at the border of groups of β-cells. (Photos provided by Arief A. Suriawinata).
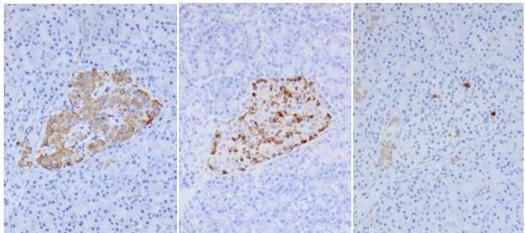
Figure 34. Triple-immunolabeling of islet hormones shows the predominance of insulin-secreting cells and their distinct distributions. This islet was stained using antibodies to insulin, glucagon and somatostatin to demonstrate beta cells (pink), alpha cells (brown), and delta cells (blue). The predominance of β-cells is obvious. α and δ-cells are typically located at the periphery of clusters β-cells. (Image provided by Vincent A. Memoli and used with permission of the American Society of Clinical Pathology).

Figure 35. Mouse and human islets stained for glucagon (Immunoperoxidase). These images show a minor species difference in the location of α-cells in mouse and human islets. In humans, α-cells appear within the islet although they seem to be on the periphery of clusters of β-cells. Compare the staining in mouse islet to Figure 36, a mouse islet stained for insulin. (Micrographs provided by Susan Bonner-Weir).
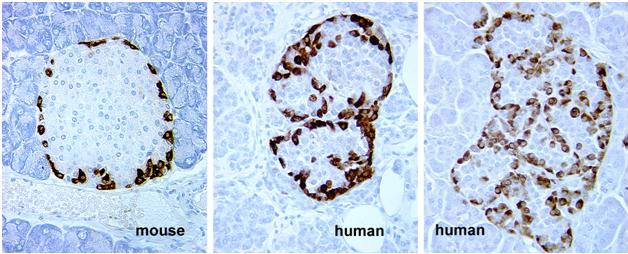
Figure 36. Mouse islet stained for insulin (Immunoperoxidase). Note that unstained cells are located in the periphery of the islet corresponding to the location of glucagon staining in Figure 35. (Micrograph provided by Susan Bonner-Weir).

Figure 37. Mouse islet stained to demonstrate pancreatic polypeptide and insulin. Immunofluorescence using antibodies to insulin (green) and neuropeptide Y (NPY) that cross‐reacts with PP (red). (Micrograph contributed by Susan Bonner‐Weir).
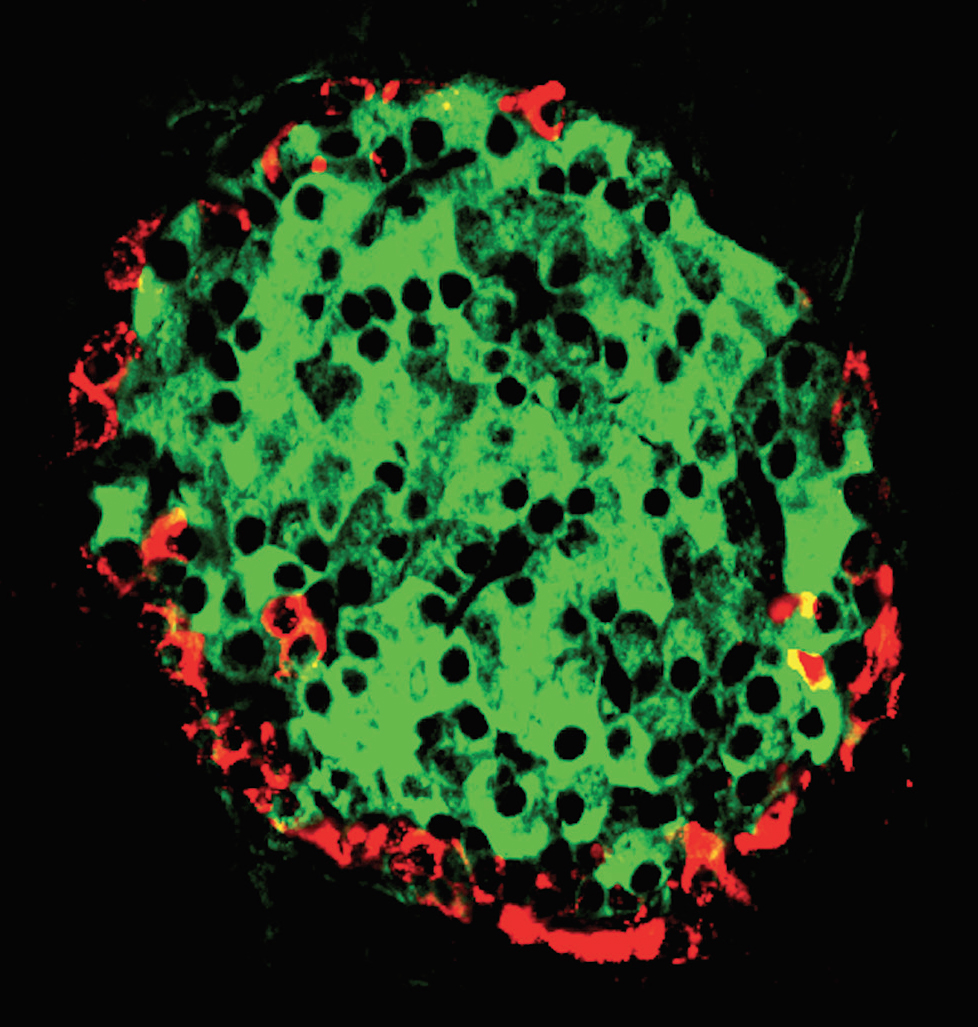
Figure 38. Human islet that was isolated during islet transplantation (Electron Micrograph). The α-, β-, and δ-cells are labeled. At the ultrastructural level, the cell types are distinguished primarily by differences in their granules. The α-cell granules are typically slightly larger than β-cell granules. δ-cell granules are typically less densely stained than the granules in α- and β-cells. At the image edge (12-1 and 9 o’clock) are interstitial (intercellular) spaces that were probably enlarged by perfusion during the islet isolation procedure. Multiple lipid vacuoles are in the cytoplasm of several islet cells--most notably in the central β-cell where lipid bodies lie at 4 and 11-12 o’clock around the nucleus. Scale bar = 4 μm. (Micrograph provided by Susan Bonner-Weir).

VIII. Online Resources
The following websites provide additional images of the pancreas. Some of the drawings are labeled in detail whereas others will challenge you to identify unlabeled structures. We recommend that you visit several of these after you review the text and images provided above. Due to the size of the files, it may take a minute or longer for some sites to open.
A drawing by Frank Netter similar to Figure 3 is posted online at: http://academicobgyn.files.wordpress.com/2010/11/pancreas-2.png&imgrefurl=http://academicobgyn.com/search/sep/feed/rss2/&h=514&w=782&sz=613&tbnid=i8zrTTDzdJwl7M:&tbnh=90&tbnw=137&zoom=1&usg=__BM8pPs5S5IKciuerloF-AgblkeE=&docid=qtye1NzwAozrdM&sa=X&ei=Rp17
{kind=link}
These two websites provide collections of drawings and photographs:
Additional images of islets:
http://en.wikipedia.org/wiki/Islets_of_Langerhans
IX. Acknowledgements
The author thanks the contributors of images listed in the text and legends; Susan Bonner-Weir for substantive input regarding the section on islets; Fred Gorelick, for preliminary review and suggestions during the preparation of the manuscript; John Williams for all Pancreapedia matters, and Jane L. Weber for editorial suggestions.
X. References
Because this is an online publication, some references will be provided as website hyperlinks when this is appropriate. Several of the references are chapters in The Pancreas: Biology, Pathobiology, and Disease. Second Edition. Edited by VLW Go et al. Raven Press Ltd. New York, 1993. These may be downloaded at <http://journals.lww.com/pancreasjournal/Pages/the-pancreas_bio_pathobio_disease.aspx>
- Barnard F.R 1921, http://www.phrases.org.uk/meanings/a-picture-is-worth-a-thousand-words.html>.
- Bockman D.E. Anatomy of the Pancreas. Chapter 1. In: The Pancreas: Biology, Pathobiology, and Disease (2nd Ed). Edited by Go VLW, et al. Raven Press Ltd., New York, pp. 1-8, 1993.
- Bockman, D.E. Morphology of the exocrine pancreas related to pancreatitis. Microsc Res Tech. 37: 509-519, 1997. PMID: 9220428
- Hellman, B. Actual distribution of the number and volume of the islets of Langerhans in different size classes in non-diabetic humans of varying ages. Nature 184: 1498-1499, 1959. PMID: 14400897
- Hellman B. The frequency distribution of the number and volume of the islets of Langerhans in man. Acta Soc Med Ups. 64: 432-460, 1959. PMID: 14400890
- Hruban R.H, Pitman M.B, Klimstra D.S. AFIP Atlas of Tumor Pathology, Fourth Series, Fascicle Tumors of the pancreas. American Registry of Pathology, Washington, DC in collaboration with the Armed Forces Institute of Pathology, Washington, DC, 2007.
- Jørgensen M.C, Ahnfelt-Rønne J, Hald J, Madsen O.D, Serup P, Hecksher-Sørensen J. An illustrated review of early pancreas development in the mouse. Endocr Rev. 28: 685-705, 2007. PMID: 17881611
- Kern H.F. Fine Structure of the Human Exocrine Pancreas. Chapter 2. In: The Pancreas: Biology, Pathobiology, and Disease (2nd Ed). Edited by Go VLW, et al. Raven Press Ltd., New York, pp. 9-19, 1993.
- Korc M. Normal Function of the Endocrine Pancreas. Chapter 38. In: The Pancreas: Biology, Pathobiology, and Disease (2nd Ed). Edited by Go VLW, et al. Raven Press Ltd., New York, pp. 751-758, 1993.
- Lee P.C and Lebenthal E. Prenatal and Postnatal Development of the Human Exocrine Pancreas. Chapter 4. In: The Pancreas: Biology, Pathobiology, and Disease (2nd Ed). Edited by Go VLW, et al. Raven Press Ltd., New York, pp. 57-73, 1993.
- Longnecker D.S and Wilson GL. Pancreas. In: Handbook of Toxicologic Pathology. Edited by Haschek-Hock WM and Rousseaux CG. Academic Press Inc., San Diego, pp. 253-278, 1991.
- Mekapogu, AR, Pothula, SP, Pirola, RC, Wilson, JS, and Apte, MV. Pancreatic Stellate Cells in Health and Disease. The Pancreapedia: Exocrine Pancreas Knowledge Base. 2020. DOI: 10.3998/panc.2020.08
- Rahier J, Wallon J, Loozen S, Lefevre A, Gepts W, HaotJ. The pancreatic polypeptide cells in the human pancreas: the effects of age and diabetes. J Clin Endocrinol Metab. 56: 441-444, 1983. PMID: 6337179
- Rahier J, Guiot Y, Goebbels R.M, Sempoux C, Henquin J.C. Pancreatic beta-cell mass in European subjects with type 2 diabetes. Diabetes Obes Metab. 10: 32-42, 2008. PMID: 18834431
- Stefan Y, Orci L, Malaisse-Lagae F, Perrelet A, Patel Y, Unger R.H. Quantitation of endocrine cell content in the pancreas of nondiabetic and diabetic humans. Diabetes. 31: 694-700, 1982. PMID: 6131002
- Valentijn K, Valentijn J.A, Jamieson J.D. Role of actin in regulated exocytosis and compensatory membrane retrieval: insights from an old acquaintance. Biochem Biophys Res Commun. 266: 652-661, 1999. PMID: 10603303
- Wittingen J, Frey C.F. Islet concentration in the head, body, tail and uncinate process of the pancreas. Ann Surg. 179: 412-414, 1974. PMID: 4593099
- Yoon K.H, Ko S.H, Cho J.H, Lee J.M, Ahn Y.B, Song K.H, Yoo S.J, Kang M.I, Cha B.Y, Lee K.W, Son H.Y, Kang S.K, Kim H.S, Lee I.K, Bonner-Weir S. Selective beta-cell loss and alpha-cell expansion in patients with type 2 diabetes mellitus in Korea. J Clin Endocrinol Metab. 88: 2300-2308, 2003. PMID: 12727989