
Entry Version:
Citation:
Pancreapedia: Exocrine Pancreas Knowledge Base, DOI: 10.3998/panc.2011.14
| Attachment | Size |
|---|---|
| 393.19 KB |
Genes: ADCY10 Human, Rat, Mouse
1. General Information
There are 10 known isoforms of adenylyl cyclase, 9 of which are membrane bound and are activated via stimulation of the Gs subunit of G-protein coupled receptors. The 10th member of this family is a soluble adenylyl cyclase, which has no trans-membrane domains. Soluble adenylyl cyclase (sAC) can be differentiated from trans-membrane AC’s by its insensitivity to forskolin and G-protein stimulation. sAC was originally isolated from the testis(3-5) and has also been found in other tissues (22, 23) . It was described as having two variants; an 187kDa full-length form (sACfl) and a 48kDa truncated version (sACt). The latter is a product of alternative splicing (13). The full-length protein consists of 2 catalytic subunits: a consensus P-loop (ATP/GTP binding site), and a leucine zipper sequence (dimerization/DNA binding domain). In contrast, sACt consists only of the 2 catalytic sub-units and is approx 20 fold more active than the full-length form (27). The catalytic domains of sAC are more closely related to those of cyanobacteria and myxobacteria than they are to trans-membrane AC’s (5, 27). Neither sACfl nor sACt contain a membrane-spanning domain. A third variant of sAC has recently been described which arises from an alternate start site preceding exon 5 and is found in somatic tissues (9). The structure of the various isoforms is detailed below in Figure 1.

Figure 1. sAC has several splice variants including one found only in somatic tissue.
As its name suggests, sAC can be found in the cytosol but has also been found associated with various sub-cellular domains including nuclei, mitochondria and microtubules (28). sAC also co-localizes with vacuolar ATPase (vATPase) subunits at the apical membrane of acid-base transporting intercalated cells in the kidney (21). The association of sAC with specific cellular organelles suggests that it can generate localized cAMP signals.
sAC was originally identified as a bicarbonate sensor (7), but can also be activated by divalent cations including Ca2+, Mg2+ and Mn2+ (16). The combination of bicarbonate and Ca2+ has been shown to synergistically activate sAC (Figure 2-A) (16). The pH sensitivity of sAC is unclear. sAC isolated from the sea urchin shows an increase in cyclase activity in the presence of manganese and increasing pH (Figure 2-B) (19). Using a mammalian sperm truncated sAC fusion protein Chen et.al. found that both basal and bicarbonate stimulated sAC activity was pH insensitive (7), while Litvin et.al. state that “. . . subtle changes in intracellular pH and/or carbon dioxide, which will be in equilibrium with bicarbonate, will result in significant changes of cellular cAMP” (16). None of these studies examines the effect of pH on the somatic form of sAC, which is different in structure form both sea urchin and mammalian sperm sAC.

Figure 2. Soluble AC is activated by calcium and bicarbonate and is pH sensitive.
cAMP derived from sAC activation has been shown to activate both PKA(29) and EPAC/RAP-1 (2, 24). sAC activation has been associated with vATPase translocation and expansion of apical microvilli (20). sAC has also been associated with CFTR activation (25, 26) , oxidative phosphorylation (1) and Na+ transport (10). Further, sAC has been reported to inhibit (15) and enhance (14) apoptosis.
2. sAC in the exocrine pancreas
In the exocrine pancreas cAMP levels increase during hyperstimulation with the secretagogue, cholecystokinin (CCK) (18). Stimulation of G-protein coupled receptors (Gs) with the ligands VIP,PACAP and secretin which activate membrane AC’s (17) or treating with membrane permeable cAMP analogs (6, 17) raises intracellular cAMP levels and results in an increase in secretagogue stimulated zymogen activation and amylase secretion. This data suggests that trans-membrane and/or soluble adenylyl cyclase play a role in acinar cell function. sAC mRNA has previously been shown to be present in the pancreas (22). We recently identified sAC in the pancreas and in pancreatic acini (Figure 3-A). We see a faint band corresponding to the truncated form of sAC in both the pancreas and in isolated acinar cells. We also found a significant band at approximately 35kD, which is not found in the testis. There is precedent for an isoform of sAC of this size in the liver (1) and in colonic epithelial cells (11). We have not seen a band corresponding to the full-length isoform of sAC in our experiments; this may be due to its absence or possibly that it is of low copy number and we are loading insufficient protein to detect this protein. Our laboratory is currently characterizing the specific isoforms present in the pancreas. As indirect evidence that sAC activity in present in pancreatic acinar cells, we have also shown that treatment with bicarbonate, a sAC activator, enhances cAMP accumulation in acinar cells (Figure 3-B).
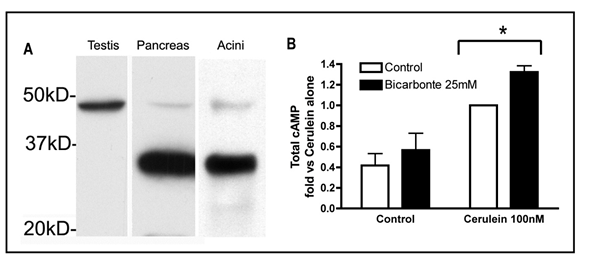
Figure 3. sAC is present in the pancreas and is activated by bicarbonate. A) Rat tissues and cells were homogenized in homogenization buffer (0.3M sucrose, 10mM HEPES, 5 mM Benzmidine and a 1X protease inhibitor cocktail from Roache). The homogenate was centrifuged 500g/10 min and PNS (supernatant) collected. 50 ug of PNS protein was loaded on a Bio-Rad Any-kD gel and transferred to PVDF. The immunoblot was probed with R21 Ab from Dr’s L. Levin and J. Buck (Cornell University School of Medicine, NYC). B) Rat pancreatic acinar cells were incubated in buffer (25 mM HEPES, (pH 7.4), 95 mM NaCl, 4.7 mM KCl, 0.6 mM MgCl2, 1 mM NaH2PO4, 10 mM glucose, 2 mM glutamine, plus 0.1% bovine serum albumin, 1× MEM-amino acids (GIBCO-BRL)). Acini were pre-incubated with the phosphodiesterase inhibitor IBMX (1 mM) for 15 min prior to a media change were acini were either put in buffer alone (control) or buffer supplemented with 25 mM sodium bicarbonate. IBMX was re-added to all wells. The experiment was initiated by the addition of cerulein (100 nM) to the appropriate wells. Control acini were incubated under room air while acini in bicarbonate-supplemented buffer were incubated under Air/C02 (95%: 5%). After 1 hour HCl was added to stop the reaction and prevent cAMP-degradation and samples were assayed using a commercial kit (cAMP direct EIA kit from Assay Designs)
3. Tools for the study of sAC
a. sAC-specific siRNA
sAC-specific siRNA can be obtained from Thermo Scientific/Dharmacon, Santa Cruz Biotechnology and a recent journal article by Li et.al. also provides information on siRNA (15). Additionally shRNA and lentivirus particle for sAC are available from Santa Cruz Biotechnology.
b. PCR Primers
PCR Primers used to differentiate between sACfl and sACt are described by Jaiswal et.al. (13).
c. Pharmacologic Inhibitors
Pharmacologic Inhibitors of sAC include 2-hydroxyestradiol, 4-hydroxyestradiol and 2-hydroxyestrone while 17-estradiol and estrone are inert and can be used as negative controls. All of these compounds are available from Steraloids Inc. KH-7 a specific inhibitor of sAC(12) and is available commercially from Tocris Biosciences and Cayman Chemical.
d. Ab’s to sAC are available from both commercial and non-commercial sources.
1. Our laboratory has been provided with several Ab’s, which work well in both rat and mouse by our collaborators Dr’s L. Levin and J. Buck (Cornell University School of Medicine, NYC).
- Ab R21 is a mouse Ab that recognizes sAC in both rat and mouse by immunoblot and in rat using indirect immunofluorescence (IF) microscopy.
- Biotinylated R21 Ab can be used for IF in mouse as well as for immunobloting of immunoprecipitated fractions.
- Ab R37 immunoprecipitates sAC from rat.
- They have also produced Ab’s to other regions of the molecule including but not limited to the N-terminus and the C-terminus.
2. Wang et.al. a group from China (26) has produced a rabbit polyclonal sAC Ab
3. Chen et.al also describe a rabbit polyclonal Ab (7)
4. Commercial sAC Ab’s are available from Prosci Incorporated and Antibodies-online.
e. sAC ELISA kits
sAC ELISA kits for human, mouse and rat are available from Antibodies-online
f. sAC activity
sAC activity can be determined in biologic samples by assaying cAMP levels in the presence and absence of a sAC inhibitor during stimulation. We have used a commercially available cAMP direct EIA kit from Assay Designs and KH7 (available from Tocris Biosciences and Cayman Chemical).
g. A knock out mouse
A knock out mouse in which exons 2-4 have been deleted has been described; this deletion leads to an almost complete inhibition of sAC dependent cAMP production in testis supernatant and spermatozoa (8). Although this mouse has little sAC activity in testis, the activity of the somatic form of sAC which starts at exon 5 is fully retained (9).
This work was supported by an RO1 grant (NIH DK54021) and a VA Merit Award to Dr. Fred Gorelick.
4. References
- Acin-Perez R, Salazar E, Kamenetsky M, Buck J, Levin LR, and Manfredi G. Cyclic AMP produced inside mitochondria regulates oxidative phosphorylation. Cell Metabolism 9: 265-276, 2009. PMID:19254571
- Branham MaT, Bustos MaA, De Blas GA, Rehmann H, Zarelli VEP, Treviño CL, Darszon A, Mayorga LS, and Tomes CN. Epac Activates the Small G Proteins Rap1 and Rab3A to Achieve Exocytosis. Journal of Biological Chemistry 284: 24825-24839, 2009. PMID:19546222
- Braun T and Dods RF. Development of a Mn-2+-sensitive, "soluble" adenylate cyclase in rat testis. Proc Natl Acad Sci U S A 72: 1097-1101, 1975. PMID:1055368
- Braun T, Frank H, Dods R, and Sepsenwol S. Mn2+-sensitive, soluble adenylate cyclase in rat testis. Differentiation from other testicular nucleotide cyclases. Biochim Biophys Acta 481: 227-235, 1977. PMID:14691
- Buck J, Sinclair ML, Schapal L, Cann MJ, and Levin LR. Cytosolic adenylyl cyclase defines a unique signaling molecule in mammals. Proc Natl Acad Sci U S A 96: 79-84, 1999. PMID:9874775
- Chaudhuri A, Kolodecik TR, and Gorelick FS. Effects of increased intracellular cAMP on carbachol-stimulated zymogen activation, secretion, and injury in the pancreatic acinar cell. American Journal of Physiology-Gastrointestinal and Liver Physiology 288: G235-G243, 2005. PMID:15458924
- Chen Y, Cann MJ, Litvin TN, Iourgenko V, Sinclair ML, Levin LR, and Buck J. Soluble adenylyl cyclase as an evolutionarily conserved bicarbonate sensor. Science 289: 625-628, 2000. PMID:10915626
- Esposito G, Jaiswal BS, Xie F, Krajnc-Franken MAM, Robben TJAA, Strik AM, Kuil C, Philipsen RLA, van Duin M, Conti M, and Gossen JA. Mice deficient for soluble adenylyl cyclase are infertile because of a severe sperm-motility defect. Proceedings of the National Academy of Sciences of the United States of America 101: 2993-2998, 2004. PMID:14976244
- Farrell J, Ramos L, Tresguerres M, Kamenetsky M, Levin LR, and Buck J. Somatic 'Soluble' Adenylyl Cyclase Isoforms Are Unaffected in Sacy^tm1Lex/Sacy^tm1Lex 'Knockout' Mice. PLoS ONE 3: e3251, 2008. PMID:18806876
- Hallows KR, Wang H, Edinger RS, Butterworth MB, Oyster NM, Li H, Buck J, Levin LR, Johnson JP, and Pastor-Soler NrM. Regulation of Epithelial Na+ Transport by Soluble Adenylyl Cyclase in Kidney Collecting Duct Cells. Journal of Biological Chemistry 284: 5774-5783, 2009. PMID:19126549
- Halm ST, Zhang J, and Halm DR. beta-Adrenergic activation of electrogenic K+ and Cl- secretion in guinea pig distal colonic epithelium proceeds via separate cAMP signaling pathways. American Journal of Physiology - Gastrointestinal & Liver Physiology 299: G81-95, 2010. PMID:20413718
- Hess KC, Jones BH, Marquez B, Chen Y, Ord TS, Kamenetsky M, Miyamoto C, Zippin JH, Kopf GS, Suarez SS, Levin LR, Williams CJ, Buck J, and Moss SB. The "Soluble" Adenylyl Cyclase in Sperm Mediates Multiple Signaling Events Required for Fertilization. Developmental Cell 9: 249-259, 2005. PMID:16054031
- Jaiswal BS and Conti M. Identification and functional analysis of splice variants of the germ cell soluble adenylyl cyclase. J Biol Chem 276: 31698-31708, 2001.
- Kumar S, Kostin S, Flacke J-P, Reusch HP, and Ladilov Y. Soluble Adenylyl Cyclase Controls Mitochondria-dependent Apoptosis in Coronary Endothelial Cells. Journal of Biological Chemistry 284: 14760-14768, 2009. PMID:11423534
- Li S, Allen KT, and Bonanno JA. Soluble adenylyl cyclase mediates bicarbonate-dependent corneal endothelial cell protection. American Journal of Physiology - Cell Physiology 300: C368-C374. PMID:21123735
- Litvin TN, Kamenetsky M, Zarifyan A, Buck J, and Levin LR. Kinetic properties of "soluble" adenylyl cyclase. Synergism between calcium and bicarbonate. J Biol Chem 278: 15922-15926, 2003. PMID:12609998
- Lu Z, Kolodecik TR, Karne S, Nyce M, and Gorelick F. Effect of ligands that increase cAMP on caerulein-induced zymogen activation in pancreatic acini. American Journal of Physiology-Gastrointestinal and Liver Physiology 285: G822-G828, 2003. PMID:12881228
- Marino CR, Leach SD, Schaefer JF, Miller LJ, and Gorelick FS. Characterization of cAMP-dependent protein kinase activation by CCK in rat pancreas. FEBS Letters 316: 48-52, 1993. PMID:7678554
- Nomura M, Beltr·n C, Darszon A, and Vacquier VD. A soluble adenylyl cyclase from sea urchin spermatozoa. Gene 353: 231-238, 2005. PMID:15978750
- Pastor-Soler N, Beaulieu Vr, Litvin TN, Da Silva N, Chen Y, Brown D, Buck J, Levin LR, and Breton S. Bicarbonate-regulated Adenylyl Cyclase (sAC) Is a Sensor That Regulates pH-dependent V-ATPase Recycling. Journal of Biological Chemistry 278: 49523-49529, 2003. PMID:14512417
- Paunescu TG, Da Silva N, Russo LM, McKee M, Lu HAJ, Breton S, and Brown D. Association of soluble adenylyl cyclase with the V-ATPase in renal epithelial cells. American Journal of Physiology - Renal Physiology 294: F130-F138, 2008. PMID:17959750
- Reed BY, Gitomer WL, Heller HJ, Hsu MC, Lemke M, Padalino P, and Pak CY. Identification and characterization of a gene with base substitutions associated with the absorptive hypercalciuria phenotype and low spinal bone density. J Clin Endocrinol Metab 87: 1476-1485, 2002. PMID:11932268
- Sinclair ML, Wang XY, Mattia M, Conti M, Buck J, Wolgemuth DJ, and Levin LR. Specific expression of soluble adenylyl cyclase in male germ cells. Mol Reprod Dev 56: 6-11, 2000. PMID:10737962
- Stessin AM, Zippin JH, Kamenetsky M, Hess KC, Buck J, and Levin LR. Soluble adenylyl cyclase mediates nerve growth factor-induced activation of Rap1. J Biol Chem 281: 17253-17258, 2006. PMID:16627466
- Sun XC, Zhai C-B, Cui M, Chen Y, Levin LR, Buck J, and Bonanno JA. HCO-3-dependent soluble adenylyl cyclase activates cystic fibrosis transmembrane conductance regulator in corneal endothelium. Am J Physiol Cell Physiol 284: C1114-1122, 2003. PMID:12519749
- Wang Y, Lam CS, Wu F, Wang W, Duan Y, and Huang P. Regulation of CFTR channels by HCO3--sensitive soluble adenylyl cyclase in human airway epithelial cells. Am J Physiol Cell Physiol 289: C1145-1151, 2005. PMID:15958523
- Wuttke MS, Buck J, and Levin LR. Bicarbonate-regulated soluble adenylyl cyclase. Jop: Journal of the Pancreas [Electronic Resource] 2: 154-158, 2001. PMID:11875252
- Zippin JH, Chen Y, Nahirney P, Kamenetsky M, Wuttke MS, Fischman DA, Levin LR, and Buck J. Compartmentalization of bicarbonate-sensitive adenylyl cyclase in distinct signaling microdomains. Faseb J 17: 82-84, 2003. PMID:12475901
- Zippin JH, Farrell J, Huron D, Kamenetsky M, Hess KC, Fischman DA, Levin LR, and Buck J. Bicarbonate-responsive "soluble" adenylyl cyclase defines a nuclear cAMP microdomain. J Cell Biol 164: 527-534, 2004. PMID:14769862