
Entry Version:
Citation:
Pancreapedia: Exocrine Pancreas Knowledge Base, DOI: 10.3998/panc.2015.1
| Attachment | Size |
|---|---|
| 456.34 KB |
Abstract
Secretory granules are located at the apex of pancreatic acinar cells. Secretagogues bind to their receptors at the basolateral membrane of acinar cells and trigger the activation of intracellular signaling pathways to elicit fusion of secretory granules with the apical plasma membrane that is followed by exocytosis of digestive pro-enzymes (the “zymogens”) into the lumen. This regulated discharge of stored macromolecules is accompanied by the secretion of solutes and water to the cell exterior to hydrate these protein-rich secretory products. Previous functional and pharmacological studies in pancreatic acinar cells and zymogen granules (ZG) had suggested that ion channels and transporters are expressed in the membrane of ZG where they contribute to maturation, fusion, exocytosis and/or fluidization of zymogens. This chapter reviews studies that have been largely published in the postgenomic era and combined biochemical, immunological, electrophysiological, pharmacological, and/or occasionally knockout methodologies to identify cloned transporters and ion channels in the membrane of ZG. Available experimental evidence indicates the presence of several ion channel and transporter proteins in ZG membranes (aquaporins, vacuolar-type H+-ATPase, zinc influx transporter SLC30A2). The evidence for the K+ channels Kv7.1 and Kir6.1, for ClC Cl- channels and the vesicular nucleotide transporter SLC17A9 in ZG is less strong. To better understand the function of these proteins in the secretory pathway further studies are needed.
1. Introductory Remarks
A review on the topic of pancreatic zymogen granule (ZG) channels and transporters and their function is timely as the last exhaustive review appeared in 2002 (107) and is manageable because of the limited number of publications that had been published in the 12 years since then. These circumstances have allowed me to carry out an in-depth and critical analysis of the published data. The advent of the post-genomic era had raised hopes that – similar to other areas of cell biology – a large number of ZG transport proteins would be identified and their role in pancreatic secretion would be elucidated. Indeed, recent studies have combined functional and molecular approaches to characterize ZG channels and transporters and their role in pancreas physiology. Yet, the fact that only a very limited number of studies have been published in this area of research is surprising as there have been tremendous developments of knowledge and methodologies available to investigate the molecular and cellular biology and physiology of the pancreas (122).
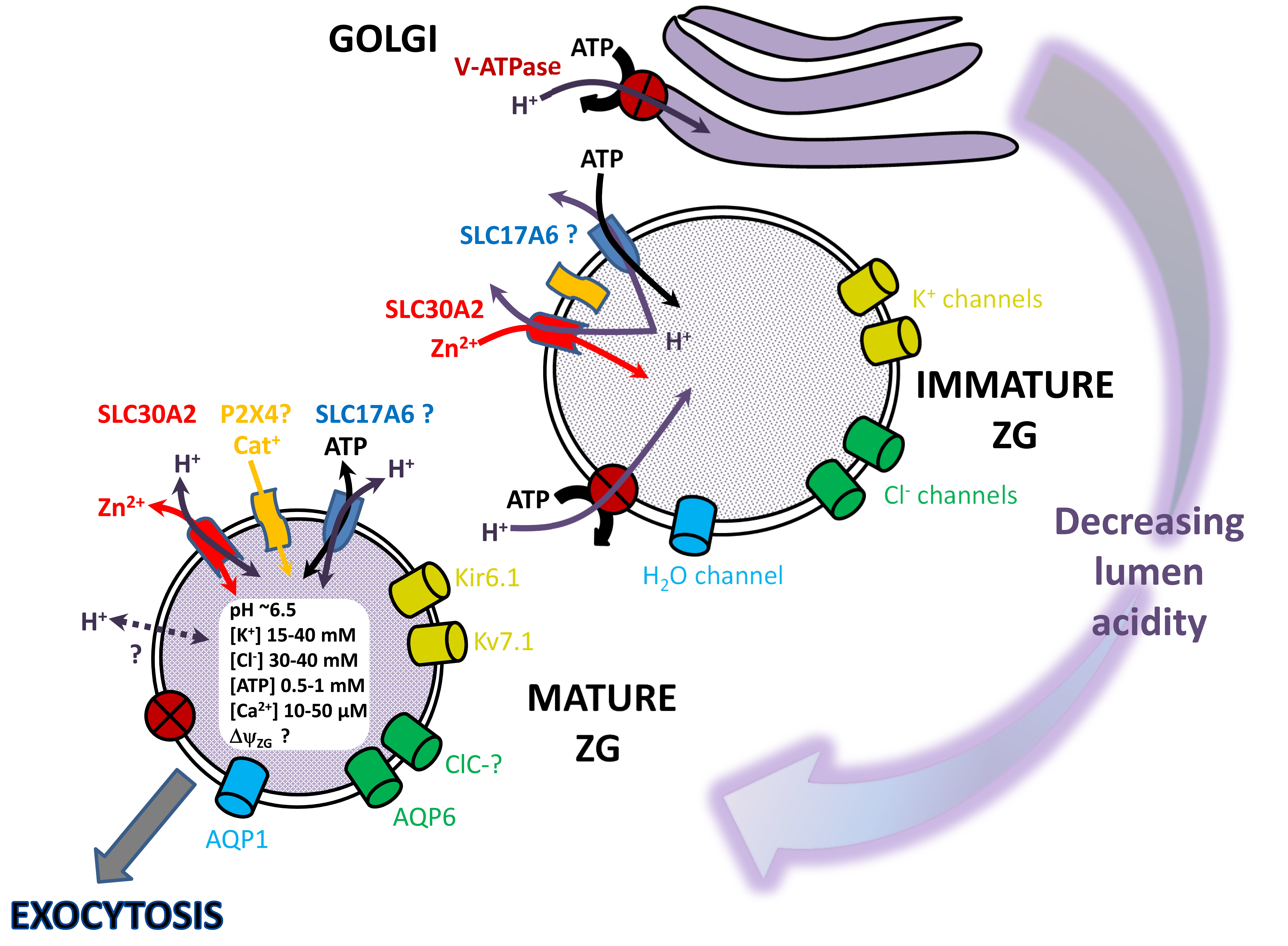
Figure 1. The model summarizes current evidence (strong or weak) for the presence of ion channels and transporters in zymogen granules (ZG) of pancreatic acinar cells. The Figure also describes relevant ion concentrations in mature ZG and emphasizes the decreasing lumen acidity during maturation along the compartments of the secretory pathway that participate in ZG maturation (for further details, see text).
Moreover, a better understanding of the physiology of pancreatic acinar cell secretion and the role played by ZG in this process seems to be a prerequisite to comprehend the pathogenesis of pancreatic disorders, such as pancreatitis, cystic fibrosis or cancer (65).
In the following paragraphs, I first review significant advances in the characterization of ZG channels and transporters in the last decade and discuss their putative role in pancreatic acinar secretion (Figure 1). The review ends on a personal assessment of possible causes for the current problems and apparent stagnation of this field of research and makes suggestions how to overcome these difficulties.
2. Proteomics
In the past, ion pathways in ZG were either characterized in permeabilized acini in which the effect of the ionic composition of the “extended cytosol” on stimulated secretion was investigated and/or by recording macroscopic ion fluxes using an osmotic swelling assay (and lysis as an end-point) in suspensions of isolated ZG (107). Such work has provided important 'background' functional information for interpreting the post-genomic work (see below).
A key advance in the last decade has been the widespread availability of proteomics that identify a large number of expressed proteins simultaneously. The use of proteomics for analysis of ZG and their membranes has propelled the identification of channels and transporters of pancreatic ZG although various caveats need to be contemplated. Several proteomic studies have been published that used highly purified ZG membranes (ZGM) to identify cloned transporters and channel proteins (9, 18, 19, 96). In most of these studies, granules were disrupted by different techniques and membranes were subjected to carbonate and/or bromide extraction, which is a standardized and reliable procedure to obtain pure membranes without peripheral proteins (14). A critical step in this isolation procedure represents the initial “purification” of ZG by either differential or continuous Percoll gradient centrifugation. These methods yield fractions that are enriched 4-8-times in α-amylase compared to homogenate, which is close to the theoretical limit of purity (77). Yet, it is practically impossible to avoid contaminations by lysosomes (96), plasma membranes (110) or membranes from other (disrupted) organelles (see electron micrographs in references (14, 110, 128)). These contaminants are, however, a major draw-back for proteomic analyses (as well as for electrophysiological studies; see the paragraph on K+ Channels) because even minimal contaminations by membranes originating from other organelles or plasma membranes may be prominent in a proteomic analysis of ZGM because of proteins that are highly expressed in these contaminating membranes. This issue needs to be considered in particular for “mitochondrial” proteins such as voltage-dependent anion channels (VDACs) and subunits of ATP-synthase that have been detected in ZGM in two independent studies (9, 19), or “plasma membrane” proteins, such as the α-subunit of Na+/K+-ATPase (96). Hence, these observations need to be taken with caution. In addition, detection of a particular channel or transporter in ZGM may not infer any physiological/functional significance of that identified protein in mature ZG (see the paragraph on Vacuolar-type H+-ATPase). Finally, it should be born in mind that several cloned channel proteins and transporters that have been identified by functional or immunological approaches have not been detected by proteomic analyses. This may be due to a relative insensitivity of proteomic approaches because of a very low expression level of these proteins in ZGM.
Interestingly, among the putative novel transporters and channels identified in ZGM by Rindler and coworkers (96) the ligand gated ATP receptor P2X4 deserves special attention because P2X4 has also been recently identified in lysosomes where it operates as a nonselective cation channel that is inhibited by luminal acidity while increasing the luminal pH in the presence of ATP causes cation channel activation (49). These functional properties in a P2X4 ATP receptor that would be located in ZG (if it is in fact present there because lysosomes easily contaminate ZGM) are attractive for ZG exocytosis considering the fact that maturing ZG lose their acidity (see the paragraph on Vacuolar-type H+-ATPase) and that the P2X4 cation channel is activated by ATP concentrations that have been measured in ZG (see paragraph on the Vesicular nucleotide transporter SLC17A9). Nonetheless, the detection of the P2X4 ATP receptor offers an example of the potentially novel information on ZG physiology that may be derived from proteomic approaches. Hence, P2X4 is a promising candidate for future studies on ZG function.
3. Channels
H2O Channels
It is a long-standing observation that isolated ZG and other secretory granules suspended in isotonic KCl buffers remain stable for hours (7, 26). Considering the packaging and condensation of a variety of osmotically active ions, small organic molecules and macromolecules in the lumen of ZG during maturation and bearing in mind that granular Cl- and K+ concentrations are lower than in the cytosol (83, 97), ZGs must exhibit low basal permeabilities for H2O, K+ and Cl- in situ and in vitro. But it has long been known that addition of cation or anion ionophores to isolated ZG suspended in iso-osmotic KCl buffers elicits granular swelling (26). The ionophore is thought to generate an electrochemical membrane potential that activates endogenous conductive pathways for the counterions and allows entry of osmotically active KCl into the granular lumen (26, 108). H2O follows and induces swelling (and lysis) implying that H2O permeable pathways, e.g. aquaporins, are present in ZGM.
The expression and function of aquaporins in pancreas has been recently reviewed (29). Aquaporin 1 (AQP1) has been identified in rat pancreatic ZGM by the full range of immunodetection methods, namely immunoblotting, immunofluorescence confocal microscopy and immunogold labeling (21). Furthermore, swelling of ZG suspended in hypo-osmotic KCl solution was detected by atomic force microscopy when 40 µM GTP was added and blocked by Hg2+, an inhibitor of AQP1, or by a functional AQP1 antibody that was raised against the carboxyl-terminal domain of AQP1. These changes of ZG swelling were paralleled by corresponding effects on fluxes of 3H2O (21). The effect of GTP on AQP1 activation is thought to be mediated by a complex of several proteins that were co-immunoprecipitated with an AQP1 antibody and included the GTP-binding protein subunit Gαi3, the inwardly rectifying K+ channel IRK8 (Kir6.1/KCNJ8), the voltage-gated Cl- channel ClC-2 and phospholipase A2 (PLA2) (1). Exposure of ZGs to either the K+ channel blocker glyburide, or the PLA2 inhibitor ONO-RS-082, blocked GTP-induced ZG swelling whereas the non-specific Cl- channel inhibitor DIDS had no effect.
Interestingly, proteomic analyses have identified AQP8, but not other AQPs, in rat ZGM (96). AQP8 may be a genuine ZG membrane protein, however a contamination by mitochondria cannot be ruled out, which express AQP8 as well (16, 61). Swelling assays of secretory granules and immunological studies have suggested that AQP5 and AQP6 mediate osmotic swelling of isolated parotid secretory granules suspended in iso-osmotic KCl solutions (72, 74), but the presence of AQP5 in parotid granules has also been contested using various immunological techniques (75).
An intriguing observation has been made by Ohta et al. (86) who found AQP12 expressed in the rough endoplasmic reticulum (rER) of rat pancreatic acinar cells. In CCK-8 treated rats, AQP12 was present in the rER and also on the membranes of ZGs near the rER. Furthermore, AQP12 knockout mice were more prone to caerulein-induced pancreatitis (86). The authors suggested that AQP12 may be involved in the mechanisms underlying the proper generation, maturation and trafficking of ZGs in the secretory pathway and that adequate H2O flux may be necessary for this process to function adequately. Altogether, it seems highly likely that AQPs (perhaps AQP1 and AQP8) are expressed in pancreatic ZGM. But generally, up to now the physiological role of AQPs in the complex process of secretory granule maturation, trafficking and exocytosis required for secretagogue-induced pancreatic acinar enzyme and fluid secretion has remained poorly understood.
K+ Channels
In pancreatic acinar cells, ZG are about 4-8-fold lower in K+ compared to the cytosol, as measured by X-ray microanalysis (83) or using a potassium-sensitive fluorescence dye (123). Hence any K+ conductive pathway should allow flux of K+ into the ZG lumen and increase its osmotic load. Based on the work by Hopfer and coworkers (32), in 1992 I proposed that (ATP- and glibenclamide-inhibitable) K+ channels are expressed in ZGM where they contribute to osmotic swelling of isolated pancreatic ZG (108). This observation was soon confirmed by others (33). Subsequent studies have suggested that glibenclamide may modulate ZG K+ permeability indirectly, possibly by binding to a 65-kDa multidrug resistance P-glycoprotein (ABCB1)-like regulatory protein (11) and/or the ZG membrane-associated protein ZG-16p (12). Additional support for the presence of K+ channels in secretory granules came from studies with mucin granules from goblet cells and suggested a Ca2+/K+ ion-exchange mechanism via parallel operation of a Ca2+ channel and a Ca2+-activated K+ channel (81). Ca2+/K+ ion-exchange has been subsequently proposed to play a critical role in a premature activation of trypsin in ZGs to promote acute pancreatitis (123). Jena and coworkers (1, 57) identified an inwardly rectifying K+ channel IRK8 (Kir6.1/KCNJ8) in ZGM by immunoblotting and measured whole vesicle currents of patched single ZGs with KCl as permeating ions that were reduced by ~15% by either 40 µM quinidine or 20 µM glibenclamide (see also the paragraph on H2O Channels). Rather, proteomic analyses of purified rat ZGM have detected the K+ channel TWIK-2/KCNK6 (96), a member of a two-pore-domain K+ channel family that produces constitutive inward rectifying K+ currents of weak amplitude and that is highly expressed in the pancreas (67, 76). However, their functional properties and their wide distribution suggest that these channels are mainly involved in the control of background K+ conductances in the plasma membranes of many cell types. Hence, it cannot be excluded that plasma membranes may have contaminated ZGM in the proteomic study reporting TWIK-2/KCNK6 expression in ZGM (96) (see also paragraph on Proteomics).
Kv7.1 (KvLQT1/KCNQ1) is a very low-conductance, voltage-gated six-membrane-spanning K+ channel distributed widely in epithelial and non-epithelial tissues, including pancreatic acinar cells (52). KCNQ1 channels associate with all five members of the KCNE β-subunit family, resulting in a β-subunit-specific change of the current characteristics (52). Kv7.1 K+ channels are blocked by the chromanol 293B (66) and the more potent and selective derivative HMR1556 (115). Indeed, we showed that 293B selectively blocks osmotic swelling of ZG induced by activation of the K+ permeability (IC50 ~10 µM) (63). Upon incorporation of ZGM into planar bilayer membranes, K+ selective channels were detected that had linear current-voltage relationships. Single channel analysis identified several K+ channel groups with distinct channel behaviors. K+ channels were inhibited by 100µM 293B or HMR1556, but not by the maxiKCa channel inhibitor, charybdotoxin (5nM). Kv7.1 protein was demonstrated by immunoperoxidase labelling of rat pancreatic tissue, immunogold labelling of ZG and immunoblotting of ZGM. 293B also inhibited cholecystokinin (CCK)-induced amylase secretion of permeabilized acini (IC50 ~10µM) when applied together with the inhibitor of a ZG nonselective cation conductance, flufenamate. Thus these data suggest that Kv7.1 accounts at least partially for ZG K+ conductance and contributes to pancreatic hormone-stimulated enzyme and fluid secretion (63).
A question of some significance for the discussion of ZG K+ conductance is whether the presence of several K+ channels with distinct biophysical properties in the bilayer experiments (63) argues against the purity of the ZGM used for the single channel studies with the planar bilayer technique. However, single patched ZG also present a mixture of channel types (57), which supports our model that ZG carry several different cation and anion conductive pathways (reviewed in (107)). To further exemplify the relevance of this concept, the effect of 293B and HMR-1556 on fluid and enzyme secretion induced by acetylcholine has also been tested in the perfused rat pancreas, but the authors were unable to observe any inhibitory effect of Kv7.1 channels blockers on secretion (60). They therefore concluded that Kv7.1 is not essential for secretagogue-mediated secretion of pancreatic acini. These results appeared to oppose the role of Kv7.1 in enzyme secretion from permeabilized rat pancreatic acini described by us in the above study (63). However, we showed that CCK-stimulated enzyme secretion in permeabilized rat pancreatic acini was abolished only if flufenamate was applied together with 293B, which is an indication that several ZG cation channels with functional redundancy are involved in enzyme secretion (63). Lastly, this experiment resolves the controversy with the study on perfused rat pancreas (60) where no effect of Kv7.1 blockers on secretagogue-induced enzyme secretion had been observed.
In a more recent study, we have followed up the role of the ZG K+ conductance in CCK-induced enzyme secretion and showed that the immunosuppressive drug cyclosporin A (CsA) (but not FK506, another commonly used immunosuppressant) activates the ZG K+ conductance and selectively increases the open probability of ZG K+ channels incorporated into planar bilayers (62). The electrophysiological (and other) data let us suggest that CsA has a direct effect on the underlying K+ channel (62). CsA also increased basal enzyme release of permeabilized rat pancreatic acini, but did not enhance CCK-induced enzyme secretion, which is in line with the results of previous studies that had investigated the role of CsA on exocrine pancreas function (41, 47) (but these authors had suggested a different mode of action of CsA). We speculated that the selective activation of ZG K+ channels by the immunosuppressant CsA may cause an increased release of digestive enzymes in situ due to osmotic swelling and lysis of ZG and that this pathological process may account for the increased incidence of post-transplant allograft pancreatitis in patients treated with CsA (but not with FK506) following hypoxia-reperfusion injury (62).
In summary, although contamination by other organelles and membranes cannot be completely excluded, it seems likely that several K+ channels with different biophysical properties and pharmacological profiles are present in ZGM. This conclusion is supported by different electrophysiological techniques (planar bilayer and single ZG patch techniques), as well as by the detection of several different K+ channel proteins by immunological or proteomic approaches. However, as for AQPs, the significance of ZG K+ channels for the physiology of secretion by pancreatic acinar cells remains unclear.
Cl- Channels
ClC Cl- channels
X-ray microanalysis studies of pancreatic acinar cells have demonstrated that the ZG Cl- concentration is about half the cytosolic Cl- concentration (83) that varies between 63 and 85 mM in resting cells when measured with a Cl- sensitive fluorescent dye (60, 129). Based on early osmotic swelling studies with isolated ZG that postulated the presence of regulated Cl- channels (reviewed in reference (107), several candidate ion channel proteins have been identified in ZGM that have been linked to ZG Cl- conductive pathways. The earliest report suggested the presence of the Cl- channel ClC-2 in ZGM (17). Using the whole-cell patch-clamp technique the authors identified a hyperpolarization-activated Cl- current in isolated pig pancreatic acinar cells. This current had the characteristic biophysical properties of ClC-2 (114) and was activated by extracellular hypotonicity, similar to ClC-2 (42), suggesting a role in volume regulation. An antiserum raised against the C-terminus of ClC-2 localized the channel to secretory granules containing amylase by immunofluorescence microscopy of acinar cells, suggesting that the channel protein incorporates into the apical plasma membrane following granule exocytosis (17). By measuring whole vesicle currents of patched single ZGs and using KCl as permeating ions, Jena and coworkers (57) detected a conductance that could be reduced by the non-selective anion transport blocker DIDS (40 µM) by ~50% and concluded that Cl- channels are present in ZGM. Immunoblots of ZG with ClC-2 and ClC-3 antibodies showed several bands, including immunoreactive bands of ~100 kDa, as expected for ClC-2 and ClC-3 (10, 106). However, ClC-2 (as well as ClC-1) is a plasma membrane Cl- channel whereas the localization of CIC-3 through ClC-7 is likely to be predominantly intracellular (105). Proteomic analyses of rat ZGM identified ClC-3 and ClC-5, thus supporting the notion that ClC channel proteins are expressed in ZGM (96). The lack of pancreas or salivary phenotype of ClC-2 (10) and ClC-3 knockout mice (5) does not exclude that ZG express these channel proteins, also because of the assumed functional redundancy of ZG ion channel proteins (107). However, immunological studies demonstrating ClC-2 and ClC-3 expression in secretory granules have to be taken with caution as long as control experiments with knockout ZG have not been performed, proving specificity of the antibodies (51). Hence, the decisive experiments proving expression of ClC proteins in ZG are still missing.
Other candidate Cl- channels
Another possible candidate for ZG anion channels may be AQP6 that operates as an anion permeable channel and not as a H2O channel because it has an asparagine residue (Asn-60) at the position corresponding to Gly-57 in AQP H2O channels (50, 68, 125). AQP6 has been detected in secretory granules of parotid acinar cells by immunological methods (73). Furthermore, osmotic swelling and lysis of secretory granules suspended in iso-osmotic KCl buffer displayed the same conductive properties as the native AQP6 channel when induced by the AQP6 activator Hg2+ (72). Interestingly, osmotic swelling showed a Hg2+-independent component (about 33%) and Hg2+-dependent swelling was about 50% DIDS-sensitive, again suggesting at least three different Cl- conductive pathways (however without functional redundancy) in these granules.
A family of putative anion channel proteins predominantly expressed in epithelial tissues that is gated by Ca2+ was named CLCA (chloride channel, Ca2+-activated). Mouse CLCA1/2 (mCLCA1/2) was detected in ZGM with a variety of immunological techniques using several antibodies that were thoroughly characterized (100, 112). Nevertheless, the gold standard of antibody characterization (namely to test the specificity of antibodies in knockout tissues used as negative controls) was not performed because mCLCA1/2 knockout mice are not available. In one study, an anion permeability was demonstrated with a ZG osmotic swelling assay whose permeability sequence, Ca2+ dependence and inhibitor sensitivity was reminiscent of CLCA associated anion currents (112). Meanwhile it has been established that hCLCA1, the human homolog of mCLCA3 that is expressed in mucous cells of the airway epithelium is a secreted protein rather than a Cl- channel (40) although it seems to modulate the conductance of endogenous Ca2+-activated Cl− channels (46). It now seems likely that all the CLCA proteins cannot act in solo as ion channels but instead interact with other proteins in a possible signaling or regulatory capacity (reviewed in (91)). Their role in ZG function is unclear and awaits further elucidation.
In summary, although there is some suggestive evidence for the expression of intracellular ClC Cl- channel proteins and for AQP6 in ZGM no strong candidate for a ZG Cl- channel has been identified. Whether a member of the TMEM16/anoctamin protein family of Ca2+-activated Cl- channels contributes to ZG anion permeability remains an interesting question (58), especially because the Ca2+-activated Cl- channel TMEM16A is expressed in the area of pancreatic acinar ZG (124).
Ca2+ Channels
Pancreatic ZG are a significant Ca2+ store (23, 83, 97) although the majority of ZG Ca2+ appears to be bound to proteins (38, 123, 124). Depending on the Ca2+ sensor used the free Ca2+ concentration in the ZG lumen varies between 9 µM (123) and 55 µM (38). How Ca2+ accumulates in ZG is unclear, but Ca2+ uptake could occur in compartments of the secretory pathway upstream of secretory granules, e.g. the rER or the Golgi apparatus, and involve active transport mediated by different Ca2+-ATPases, such as SERCA (Sarcoplasmic/endoplasmic Reticulum Calcium ATPase) and SPCA (Secretory Pathway Calcium ATPase) pumps (120). Ca2+ released from ZG into the pancreatic juice may have a paracrine function on duct cells that express Ca2+-sensing receptors on their apical surface, as proposed by Bruce et al. (13).
Secretagogues have been shown to evoke local increases of cytosolic Ca2+ in the secretory pole region of pancreatic acinar cells (116). Following studies demonstrating IP3-induced Ca2+ release from acidic neuroendocrine chromaffin granules induced by the second messenger inositol 1,4,5-trisphosphate (IP3), experimental evidence has suggested that secretory granules from both endocrine and exocrine cells, including pancreatic acinar and airway goblet cells, are IP3-sensitive Ca2+ stores that play a significant role in Ca2+-dependent secretion (reviewed in (93, 126). In particular, using ZG isolated by differential centrifugation or intact pancreatic acinar cells, Petersen, Gerasimenko and colleagues have demonstrated release of Ca2+ from an acidic store in the apical pole of acinar cells induced by IP3, cyclic-ADP ribose or nicotinic acid adenine dinucleotide phosphate (i.e. suggestive of functional ryanodine receptors) that they associated with ZG (37, 38, 117). Gerasimenko and coworkers (35, 36) (reviewed in (93)) subsequently hinted at the importance of this acidic ZG store for the pathogenesis of alcohol-induced pancreatitis. However, two independent studies have challenged these observations in pancreatic (128) and parotid acinar cells (80). These authors provided functional and biochemical evidence indicating that ZGM do not express IP3 or ryanodine receptors provided that ZG are not contaminated by other organellar structures, especially ER and mitochondria. Although the study of Gerasimenko et al. (38) had shown a three-fold enrichment of the ZG marker α-amylase in their ZG preparation the assay used to exclude ER contamination relied on the fluorescent dye DiOC6(3) [sic!] that is normally applied to assess ΔΨ of energized mitochondria (56). Using the ZG isolation procedure of Gerasimenko et al. (38), Yule et al. (128) showed disrupted mitochondria and ER membranes contaminating ZG by electron microscopy. Interestingly, Gerasimenko et al. (34) have recently revisited this issue in a 2014 review where they conceded that the main apical Ca2+ pool that is responsible for IP3-induced Ca2+ release may represent an acidic ER store. This conclusion may be more accurate than the earlier association of acidic stores with ZG and appears to be supported by three lines of evidence: 1) The majority of ZG are not acidic (87); 2) ER also co-localizes with ZG (39, 119); 3) an acidic IP3-sensitive ER Ca2+ pool has been previously demonstrated that may operate via parallel operation of a vacuolar-type H+ pump and a Ca2+/H+ exchanger, both in pancreatic (109, 111) and parotid acinar cells (113).
From the available evidence, it appears now very likely that ZG are not an IP3 sensitive Ca2+ pool because IP3 receptors are not expressed in ZGM. It remains an open question whether other modes of Ca2+ release mechanisms are operative in ZG. Interestingly, Hille and coworkers (30) have recently shown a release of Ca2+ via “store-operated” Orai channels in the membrane of secretory granules from neuroendocrine PC12 cells (they did not detect IP3 receptors in these secretory granules, in contrast to previous studies (126)). These granule Orai channels are activated by regulator stromal interaction molecule 1 (STIM1) on the ER and may serve to raise local cytoplasmic Ca2+ concentrations for refilling of Ca2+ stores of the ER and to promote exocytosis (30).
4. Transporters
Vacuolar-type H+-ATPase
One of the hallmarks of many vesicular compartments is the expression of a vacuolar-type H+-ATPase (V-ATPase) that acts to acidify the vesicular lumen (31). Subunits of the V-ATPase have been clearly identified in the membrane of rat pancreas ZG by immunological methods (99) and by proteomic analyses (19, 96). Nevertheless, conflicting results were obtained in several early studies when measuring lumenal pH of secretory granules in freshly isolated acini or purified granules: Using fluorescence microscopy and the weak base acridine orange (AO) that accumulates in acidic organelles and changes its fluorescence from green to orange when concentrated, ZG were found to be acidic whereas parotid secretory granules showed a neutral pH (6, 7, 28, 82). In a landmark study, Orci et al. (87) used the probe 3-(2,4-dinitroanilino)-3-amino-N-methyldipro-pylamine (DAMP), a weak base that accumulates in the lumen of acidic organelles, where it can be fixed (4). Quantitative immunostaining of DAMP in conjunction with electron microscopy was used to measure pH in the secretory pathway of fixed isolated pancreatic acini. The data revealed that condensing vacuoles are acidic but lose their acidity during maturation (87). To account for these results, the authors speculated that the loss of acidity of maturing ZG may be the consequence of either removal of H+-pump subunits, inactivation of the V-ATPase or closing of an anion conductance. It is noteworthy that it took more than 20 years to confirm the observation that the acidity of ZG decreases during maturation in freshly isolated pancreas acini using AO and live-cell two-photon imaging (8), thus suggesting that earlier AO measurements with fluorescence microscopic techniques were not adequate or insensitive. The study by Thorn and coworkers (8) also showed that the H+ concentration of mature granules is not affected by pre-incubation with a blocker of the V-ATPase and that upon secretagogue-stimulated secretion the pH in the acinar lumen never drops below 7.0 which is an indication that ZG pH is at the most weakly acidic. A decrease of acidity in the secretory pathway during maturation is anomalous as in most secretory organelles along the secretory pathway the luminal pH drops from the near-cytosolic value of 7.2 within the ER to 6.7–6.0 along the Golgi complex to approximately 5.2–5.7 in secretory granules and is often associated with a reciprocal decrease of a H+ permeability (see (90) for review). This is particularly the case in neuroendocrine cells where the increasing electrochemical gradient H+ gradient generated by the V-ATPase is also used to store cargo (e.g. neurotransmitters or metal ions) above equilibrium with the cytosol (31, 54). In contrast, mature pancreatic ZGs do not store ATP (45) or Zn2+ (83) above cytosolic concentrations, which is in accordance with a reduced acidity of mature ZG. How ZG (as well as parotid secretory granules; see (6, 7) lose their acidity during maturation remains to be investigated, but this could be the consequence of a differential activity or expression of V-ATPases and/or differences in H+ permeabilities (90), or activation of exchange pathways for counterions, e.g. Na+/H+-, Ca2+/H+- or Cl-/H+-exchangers (2, 3, 88, 105) along the secretory pathway of pancreatic (and parotid) acinar cells that would result in a dissipation of the pH gradient during maturation.
Vesicular nucleotide transporter SLC17A9
SLC17A9 (also named VNUT for Vesicular Nucleotide Transporter) is a vesicular ATP transporter widely expressed in various organs, but predominantly in the adrenal gland, brain, and thyroid gland (pancreas was not tested) (101). SLC17A9 may mediate concentrative accumulation of ATP in secretory vesicles, such as chromaffin or synaptic vesicles, to a luminal concentration of 100 mM or more (reviewed in references (15, 53). SLC17A9 protein shows a relative mobility varying between 61 and 68 kDa in SDS-PAGE depending on the origin of the protein (native or recombinant). When reconstituted into proteoliposomes, SLC17A9 transports ATP (as well as ADP, GTP and UTP as shown by Cis inhibition experiments) and uses a ΔΨ (positive inside) but not a ΔpH (the vesicle lumen being acidic) as driving force for uptake. ATP transport depends on external Cl- and saturates at ~4 mM external Cl-. ATP uptake is inhibited by 4,4-diisothiocyanatostilbene-2,2-disulfonate (DIDS) with an IC50 of ~1.5 µM. Evans blue is even more potent (IC50 ~40 nM) whereas 200 µM atractyloside inhibits SLC17A9 only in the presence of Mg2+. All these properties are similar to those that have been reported for ATP uptake in chromaffin granules and granule membrane ghosts (53). Although ATP transport rate seemed to be independent of Ca2+ or Mg2+, subsequent studies by the same group showed that SLC17A9 also transports both divalent cations in an ATP- and Cl-- dependent manner and is driven by an inside-positive ΔΨ, but not by an inside-positive ΔpH, and with a similar inhibitor sensitivity as ATP transport (79). Hence, divalent cation transport probably occurs by complexation with ATP and may contribute to vesicular accumulation of divalent cations in secretory vesicles as well.
It has been hypothesized that during pancreas secretion evoked by secretagogues ATP is released from ZG into the primary fluid to mediate paracrine signaling between acinar and duct cells and thereby coordinates ductal and acinar fluid secretion (84). This may occur via purinergic receptors expressed on the apical membrane of duct cells (70). Indeed, pancreatic ZG do contain ATP: From measurements with freshly isolated ZG it has been estimated that the in vivo concentration of ATP in ZG is 0.5-1 mM ATP (45), a more than 100-times lower concentration than in neuroendocrine secretory vesicles expressing SLC17A9 and a concentration that is also below the average cytosolic concentration of 2-5 mM ATP (15), thus making concentrative ATP transport unnecessary and therefore unlikely. Nevertheless, Novak and coworkers have suggested that SLC17A9 is expressed in the membrane of ZG where it mediates ATP uptake (44, 45). Their experimental evidence supports expression of SLC17A9 in ZGM, e.g. detection of a ca. 65 kDa protein with a SLC17A9-specific antiserum, co-localization of immunolabeling for SLC17A9 with the ZG membrane marker Rab3D, or inhibition of ATP uptake into ZG by 100 µM DIDS and Evans blue (44, 45). Furthermore, induction of rat AR42J acinar cells into a secretory phenotype by dexamethasone administration increases carbachol-induced ATP release and SLC17A9 expression. In Haanes’ and Novak’s work in isolated ZG, ATP uptake depends on external Mg2+ and high Cl-, is driven by an inside-positive ΔpH (either generated by an artificial pH gradient or as demonstrated by bafilomycin A1-sensitive uptake) as well as an inside-positive K+ diffusion potential (although it is not clear whether this K+ diffusion potential is physiologically significant) and shows no Cis inhibition by ADP or UTP (45), all properties which differ from the transport properties of SLC17A9. It may be argued that results obtained in intact ZG may differ from transport of a protein reconstituted into proteoliposomes (45). However, transport of reconstituted SLC17A9 matches transport in intact chromaffin granules (53, 101). Hence, ATP uptake and expression of SLC17A9 in ZG could be observations that are not causally linked. Experiments providing evidence for a causal relationship between ZG ATP accumulation and SLC17A9 are required to confirm the postulated role of SLC17A9 in (concentrative?) ATP uptake into ZG, e.g. by knocking down the transporter and determining ZG ATP concentration and/or secretagogue-induced ATP release from AR42J cells. Moreover, it remains to be tested whether SLC17A9 (if it is indeed expressed in ZGM; another member of this transporter family could be expressed) may be involved in the accumulation of Ca2+ in ZG that, apart from the endoplasmic reticulum represents a significant intracellular Ca2+ pool (23, 97, 98). In summary, the available evidence that SLC17A9 is responsible for concentrative ATP uptake into pancreatic ZG is weak.
Zinc transporter SLC30A2
The SLC30A2 gene encodes a zinc (Zn2+) transporter (also named ZnT2 for Zinc Transporter 2) that was cloned from a rat kidney cDNA expression library by complementation of a Zn2+-sensitive Baby Hamster Kidney cell line (89) and that is also highly expressed in pancreas (reviewed in (48, 102)). Using GFP-ZnT2 fusion proteins and immunofluorescence studies, the transporter was found to be localized in vesicular membranes of acidic intracellular compartments (as opposed to the Zn2+ efflux transporter SLC30A1/ZnT1 that is expressed in the plasma membrane). The authors suggested that ZnT2 contributes to protection against Zn2+ toxicity by facilitating Zn2+ influx into an endosomal/lysosomal compartment thereby lowering cytosolic Zn2+ jointly with ZnT1 (89). Subsequent studies have revealed that, in addition to the pancreas, SLC30A2 is also expressed in mammary gland, prostate, retina, small intestine and kidney where it is localized on the membrane of intracellular organelles (e.g. endosomal/lysosomal and secretory vesicles) and sequesters cytoplasmic Zn2+ for secretion, storage, or for use in proteins that require Zn2+ for their activities (reviewed in (48, 102)). As an example, mutation or knockdown of SLC30A2 results in reduced Zn2+ secretion from mammary gland epithelial cells in vivo and in vitro (22). The transport mechanism of SLC30A2 has not been yet elucidated, yet several studies indicate that vesicular Zn2+ transport mediated by mammalian SLC30 transporters (SLC30A1 and SLC30A5) is catalyzed by H+/Zn2+ exchange (85, 103), and for SLC30A5 may be driven by the vesicular H+ gradient generated by V-ATPases (85).
In pancreatic islet β-cells granules Zn2+ is almost 40-times enriched compared to the cytosol whereas no difference between cytosolic and acinar ZG Zn2+ concentration was found, as measured by X-ray microanalysis (83). In β-cells granules (and other neuroendocrine secretory granules) SLC30A8 has been identified as the Zn2+ transporter that is responsible for insulin maturation and crystallization before secretion (20), and its possible importance in the etiology of diabetes has been highlighted (64). Because neuroendocrine secretory granules are acidic (54, 107) SLC30A8-mediated Zn2+ accumulation against a concentration gradient is likely driven by the vesicular H+ gradient generated by vacuolar-type H+-ATPases and could occur via H+/Zn2+ exchange, as demonstrated for SLC30A1 and SLC30A5 (85, 103).
The situation in ZG is much less clear. In rats, Zn2+ deficiency caused Zn2+ depletion in acinar but not in β-cells (104), which is in line with the ability of β-cells to store Zn2+ in insulin granules in a slowly-exchanging intra-granule compartment and also with the apparent inability of pancreatic ZG to enrich Zn2+ (83) (although free Zn2+ concentration in the granular compartment could exceed free cytosolic Zn2+ concentration considering the differences in affinities of Zn2+-binding proteins in cytosol [e.g. Zn2+-metallothionein with nano- to picomolar stability constants] versus granular compartment [e.g. zymogens with micromolar stability constants]). Liuzzi et al. (69) showed that Zn2+ depletion in mice is associated with a reduction of pancreatic acinar SLC30A1 and SLC30A2, and the authors speculated that SLC30A1/2 may mediate lowering of cytosolic Zn2+ in pancreatic acinar cells of Zn2+ replete animals. Based on a study showing secretion of the cytosolic Zn2+-binding protein metallothionein into pancreatic juice (27), De Lisle and coworkers suggested that exocytotic secretion of Zn2+ by pancreatic acinar cells could contribute to physiological Zn2+ homeostasis by controlling Zn2+ losses into the pancreatic fluid (24, 69). Recently, Cousins and coworkers showed SLC30A2 expression in ZG, and reduction of the Zn2+ diet of mice was paralleled by decreases of Zn2+ concentrations and SLC30A2 expression in ZG (43). Rat AR42J acinar cells when induced into a secretory phenotype with dexamethasone exhibited increased SLC30A2 expression that was associated with a reduction of cytosolic 65Zn2+ content and an increase of the 65Zn2+ content of ZG. SLC30A2 knockdown in the AR42J cells using siRNA (~70% mRNA reduction) increased cytoplasmic 65Zn2+ by 36% and decreased ZG 65Zn2+ by 15%, suggesting that SLC30A2 mediates the sequestration of Zn2+ into ZG (43). The relatively low impact of SLC30A2 knockdown on ZG Zn2+ content may be related to the observation that ZG do not accumulate Zn2+ above cytosolic concentration (which is probably caused by the loss of acidity of ZG during maturation; see the paragraph on Vacuolar-type H+-ATPase), or because other Zn2+ transporters may more significantly contribute to Zn2+ flux across the ZG membrane. What could be the significance of the presence of Zn2+ in ZG? For instance, it is known that ZG contain Zn2+-metalloenzymes, such as carboxypeptidases that require Zn2+ for their function (118), but also for inhibition of their function at micromolar Zn2+ concentrations (59). Although SLC30A2 is expressed in ZG, Zn2+ uptake via SLC30A2 (or other members of the SLC30 protein family) is more likely to be operative in acidic organelles of the secretory pathway (e.g. the Golgi apparatus or immature granules) during or immediately after protein synthesis (92). Whether secretagogue-evoked exocytotic secretion of Zn2+ from ZG (43) and/or secretion of cytosolic Zn2+-metallothionein by a mechanism independent of ZG exocytosis (27) are effective means to regulate Zn2+ body homeostasis remains to be shown. In fact, a regulation of the expression of transporters for Zn2+ uptake (from dietary sources and/or the pancreatic juice) in the intestinal mucosa may be a more effective and sensitive mechanism to control Zn2+ body homeostasis (25, 55, 71); however see a divergent view in (130)).
5. Conclusion: Problems, Questions, Outlook
From a scientific perspective, the major unresolved issue so far is not so much the presence or absence of specific ZG channels and transporters (there is no doubt that channels and transporters are expressed in the membrane of ZG) but rather to understand their physiological role in the process of fusion and exocytosis. Here the development of functional channel/transporter knockout animal models is mandatory to move the field forward. In this context, the anomalous property of the pancreatic acinar cell displaying a decreasing acidity of maturing ZG (Figure 1) remains an enigma because other secretory cells, in particular neuroendocrine cells, exhibit an increasing acidity along the secretory pathway (90). This is particularly puzzling because very recent studies in neuroendocrine cells have highlighted the important role of the V0 domain of the V-ATPase as a sensor of intragranular pH that identifies and regulates the ability of granules to undergo exocytosis (95, 121). Hence, what is the significance of a decreasing acidity of maturing ZG?
More generally, during the writing of this review several questions arose: Firstly, why is there so little understanding of the function of ZG channels and transporters in pancreatic acinar secretion despite an increasing number of identified ZG transporter and channel proteins? Not surprisingly, substantial molecular evidence for channels and transporters in ZG contrasts with the “bag-like” models found in influential reviews on the topic (e.g. (78)). Secondly, why are there so few studies linking molecular data with function? In other words, there appears to be a large gap between functional and expression studies. Thirdly and most importantly, why is there a lack of (functional) channel/transporter knockout animal models (despite substantial progress in these methodologies) on which the validity of dominant physiological and pathophysiological concepts could be vigorously tested? Finally, why are the dominant models of pancreatic acinar physiology and pathophysiology almost exclusively based on functional studies in cells and with a focus on Ca2+ signaling (93, 94)? Quite revealingly, these models ironically also reproduce the oscillating relevance of ZG as a physiological IP3-sensitive Ca2+ store that may (or may not) be involved in diseases, such as pancreatitis (see paragraph on Ca2+ Channels; for instance compare (34) with (37, 38)). Despite the undisputed relevance of Ca2+ signaling for the physiology of acinar cell secretion (see the special issue in Cell Calcium on “Ca2+ signaling and secretory function” (127)), this focus may be too narrow to describe the pathophysiology of pancreatitis (especially when these studies have been performed on isolated acinar cells (see for instance (36)) whose relevance as a model of pancreatitis is debatable). In this author’s view, it is to be feared that a lack of plurality of working hypotheses and methodologies will prevent further generation of knowledge and actually hinder further progress in understanding the physiology of pancreatic acinar cells and the role of ZG. This strategy also represents a hazard for current translational research and medical applications as significant gaps of understanding of pancreatic acinar physiology still prevail although currently largely ignored.
Hopefully, a future generation of researchers in the area of pancreatic acinar physiology will overcome current difficulties in areas of research outside of the current dominating subjects. As this review has sought to demonstrate, many unresolved – and neglected – problems remain in ZG physiology. It is likely that they also require attention before we can frame more accurate concepts of efficient preventive and therapeutic strategies for diseases of the pancreas, such as acute and chronic pancreatitis.
Acknowledgements
I thank Prof. John A. Williams for encouraging me to write this review. I am grateful to my collaborator Jun.-Prof. Wing-Kee Lee for relentless discussions and her outstanding expertise in the planar bilayer technique. I also thank my former graduate student, Assistant Professor Matthias Braun (http://www.pmcol.ualberta.ca/people/Faculty/BraunDetail.html) for contributing to several important studies on K+ channels in zymogen granules. Dr. Austin C. Elliott (University of Manchester, U.K.) critically read the manuscript and provided many valuable suggestions. Prof. Wolfgang Maret (King’s College, London, U.K.) drew my attention to the significance of different cellular zinc pools and buffers for cellular zinc homeostasis. Financial support for my studies on zymogen granules was made possible by funds from the Deutsche Mukoviszidose e.V. and the University of Witten/Herdecke.
6. References
- Abu-Hamdah R, Cho WJ, Cho SJ, Jeremic A, Kelly M, Ilie AE, and Jena BP. Regulation of the water channel aquaporin-1: isolation and reconstitution of the regulatory complex. Cell Biol Int 28: 7-17, 2004. PMID: 14759764
- Anderie I, Blum R, Haase W, Grinstein S, and Thévenod F. Expression of NHE1 and NHE4 in rat pancreatic zymogen granule membranes. Biochem Biophys Res Commun 246: 330-336, 1998. PMID: 9610358
- Anderie I and Thévenod F. Evidence for involvement of a zymogen granule Na+/H+ exchanger in enzyme secretion from rat pancreatic acinar cells. J Membr Biol 152: 195-205, 1996. PMID: 8672081
- Anderson RG and Pathak RK. Vesicles and cisternae in the trans Golgi apparatus of human fibroblasts are acidic compartments. Cell 40: 635-643, 1985. PMID: 3882239
- Arreola J, Begenisich T, Nehrke K, Nguyen HV, Park K, Richardson L, Yang B, Schutte BC, Lamb FS, and Melvin JE. Secretion and cell volume regulation by salivary acinar cells from mice lacking expression of the Clcn3 Cl- channel gene. J Physiol 545: 207-216, 2002. PMID: 12433961
- Arvan P and Castle JD. Isolated secretion granules from parotid glands of chronically stimulated rats possess an alkaline internal pH and inward-directed H+ pump activity. J Cell Biol 103: 1257-1267, 1986. PMID: 3021780
- Arvan P, Rudnick G, and Castle JD. Osmotic properties and internal pH of isolated rat parotid secretory granules. J Biol Chem 259: 13567-13572, 1984. PMID: 6490664
- Behrendorff N, Floetenmeyer M, Schwiening C, and Thorn P. Protons released during pancreatic acinar cell secretion acidify the lumen and contribute to pancreatitis in mice. Gastroenterology 139: 1711-1720, 1720 e1711-1715, 2010. PMID: 20691184
- Berkane AA, Nguyen HT, Tranchida F, Waheed AA, Deyris V, Tchiakpe L, Fasano C, Nicoletti C, Desseaux V, Ajandouz el H, Comeau D, Comeau L, and Hiol A. Proteomic of lipid rafts in the exocrine pancreas from diet-induced obese rats. Biochem Biophys Res Commun 355: 813-819, 2007. PMID: 17320817
- Bosl MR, Stein V, Hubner C, Zdebik AA, Jordt SE, Mukhopadhyay AK, Davidoff MS, Holstein AF, and Jentsch TJ. Male germ cells and photoreceptors, both dependent on close cell-cell interactions, degenerate upon ClC-2 Cl- channel disruption. EMBO J 20: 1289-1299, 2001. PMID: 11250895
- Braun M, Anderie I, and Thévenod F. Evidence for a 65 kDa sulfonylurea receptor in rat pancreatic zymogen granule membranes. FEBS Lett 411: 255-259, 1997. PMID: 9271216
- Braun M and Thévenod F. Photoaffinity labeling and purification of ZG-16p, a high-affinity dihydropyridine binding protein of rat pancreatic zymogen granule membranes that regulates a K+-selective conductance. Mol Pharmacol 57: 308-316, 2000. PMID: 10648640
- Bruce JI, Yang X, Ferguson CJ, Elliott AC, Steward MC, Case RM, and Riccardi D. Molecular and functional identification of a Ca2+ (polyvalent cation)-sensing receptor in rat pancreas. J Biol Chem 274: 20561-20568, 1999. PMID: 10400686
- Burnham DB, Munowitz P, Thorn N, and Williams JA. Protein kinase activity associated with pancreatic zymogen granules. Biochem J 227: 743-751, 1985. PMID: 4004796
- Burnstock G. Physiology and pathophysiology of purinergic neurotransmission. Physiol Rev 87: 659-797, 2007. PMID: 17429044
- Calamita G, Ferri D, Gena P, Liquori GE, Cavalier A, Thomas D, and Svelto M. The inner mitochondrial membrane has aquaporin-8 water channels and is highly permeable to water. J Biol Chem 280: 17149-17153, 2005. PMID: 15749715
- Carew MA and Thorn P. Identification of ClC-2-like chloride currents in pig pancreatic acinar cells. Pflugers Arch 433: 84-90, 1996. PMID: 9019735
- Chen X, Ulintz PJ, Simon ES, Williams JA, and Andrews PC. Global topology analysis of pancreatic zymogen granule membrane proteins. Mol Cell Proteomics 7: 2323-2336, 2008. PMID: 18682380
- Chen X, Walker AK, Strahler JR, Simon ES, Tomanicek-Volk SL, Nelson BB, Hurley MC, Ernst SA, Williams JA, and Andrews PC. Organellar proteomics: analysis of pancreatic zymogen granule membranes. Mol Cell Proteomics 5: 306-312, 2006. PMID: 16278343
- Chimienti F, Devergnas S, Favier A, and Seve M. Identification and cloning of a beta-cell-specific zinc transporter, ZnT-8, localized into insulin secretory granules. Diabetes 53: 2330-2337, 2004. PMID: 15331542
- Cho SJ, Sattar AK, Jeong EH, Satchi M, Cho JA, Dash S, Mayes MS, Stromer MH, and Jena BP. Aquaporin 1 regulates GTP-induced rapid gating of water in secretory vesicles. Proc Natl Acad Sci USA 99: 4720-4724, 2002. PMID: 11917120
- Chowanadisai W, Lonnerdal B, and Kelleher SL. Identification of a mutation in SLC30A2 (ZnT-2) in women with low milk zinc concentration that results in transient neonatal zinc deficiency. J Biol Chem 281: 39699-39707, 2006. PMID: 17065149
- Clemente F and Meldolesi J. Calcium and pancreatic secretion. I. Subcellular distribution of calcium and magnesium in the exocrine pancreas of the guinea pig. J Cell Biol 65: 88-102, 1975. PMID: 165206
- Cousins RJ. Gastrointestinal factors influencing zinc absorption and homeostasis. Int J Vitamin Nutr Res 80: 243-248, 2010. PMID: 21462106
- Cragg PJ. Artificial transmembrane channels for sodium and potassium. Sci Prog 85(Pt 3): 219-241, 2002. PMID: 12449537.
- De Lisle RC and Hopfer U. Electrolyte permeabilities of pancreatic zymogen granules: implications for pancreatic secretion. Am J Physiol Gastrointest Liver Physiol 250: G489-G496, 1986. PMID: 3754390
- De Lisle RC, Sarras MP, Jr., Hidalgo J, and Andrews GK. Metallothionein is a component of exocrine pancreas secretion: implications for zinc homeostasis. Am J Physiol Cell Physiol 271: C1103-1110, 1996. PMID: 8897816
- De Lisle RC and Williams JA. Zymogen granule acidity is not required for stimulated pancreatic protein secretion. Am J Physiol 253: G711-719, 1987. PMID: 2447787
- Delporte C. Aquaporins in salivary glands and pancreas. Biochim Biophys Acta 1840: 1524-1532, 2014. PMID: 23954206
- Dickson EJ, Duman JG, Moody MW, Chen L, and Hille B. Orai-STIM-mediated Ca2+ release from secretory granules revealed by a targeted Ca2+ and pH probe. Proc Natl Acad Sci USA109: E3539-3548, 2012. PMID: 23184982
- Forgac M. Vacuolar ATPases: rotary proton pumps in physiology and pathophysiology. Nat Rev Mol Cell Biol 8: 917-929, 2007. PMID: 17912264
- Gasser KW, DiDomenico J, and Hopfer U. Potassium transport by pancreatic and parotid zymogen granule membranes. Am J Physiol 255: C705-711, 1988. PMID: 3202144
- Gasser KW and Holda JR. ATP-sensitive potassium transport by pancreatic secretory granule membrane. Am J Physiol 264: G137-142, 1993. PMID: 8430796
- Gerasimenko J, Peng S, and Gerasimenko O. Role of acidic stores in secretory epithelia. Cell Calcium 55: 346-354, 2014. PMID: 24832105
- Gerasimenko JV, Lur G, Ferdek P, Sherwood MW, Ebisui E, Tepikin AV, Mikoshiba K, Petersen OH, and Gerasimenko OV. Calmodulin protects against alcohol-induced pancreatic trypsinogen activation elicited via Ca2+ release through IP3 receptors. Proc Natl Acad Sci U S A 108: 5873-5878, 2011. PMID: 21436055
- Gerasimenko JV, Lur G, Sherwood MW, Ebisui E, Tepikin AV, Mikoshiba K, Gerasimenko OV, and Petersen OH. Pancreatic protease activation by alcohol metabolite depends on Ca2+ release via acid store IP3 receptors. Proc Natl Acad Sci U S A 106: 10758-10763, 2009. PMID: 19528657
- Gerasimenko JV, Sherwood M, Tepikin AV, Petersen OH, and Gerasimenko OV. NAADP, cADPR and IP3 all release Ca2+ from the endoplasmic reticulum and an acidic store in the secretory granule area. J Cell Sci 119: 226-238, 2006. PMID: 16410548
- Gerasimenko OV, Gerasimenko JV, Belan PV, and Petersen OH. Inositol trisphosphate and cyclic ADP-ribose-mediated release of Ca2+ from single isolated pancreatic zymogen granules. Cell 84: 473-480, 1996. PMID: 8608601
- Gerasimenko OV, Gerasimenko JV, Rizzuto RR, Treiman M, Tepikin AV, and Petersen OH. The distribution of the endoplasmic reticulum in living pancreatic acinar cells. Cell Calcium 32: 261-268, 2002. PMID: 12543088
- Gibson A, Lewis AP, Affleck K, Aitken AJ, Meldrum E, and Thompson N. hCLCA1 and mCLCA3 are secreted non-integral membrane proteins and therefore are not ion channels. J Biol Chem 280: 27205-27212, 2005. PMID: 15919655
- Groblewski GE, Wagner AC, and Williams JA. Cyclosporin A inhibits Ca2+/calmodulin-dependent protein phosphatase and secretion in pancreatic acinar cells. J Biol Chem 269: 15111-15117, 1994. PMID: 7515049
- Grunder S, Thiemann A, Pusch M, and Jentsch TJ. Regions involved in the opening of CIC-2 chloride channel by voltage and cell volume. Nature 360: 759-762, 1992. PMID: 1334533
- Guo L, Lichten LA, Ryu MS, Liuzzi JP, Wang F, and Cousins RJ. STAT5-glucocorticoid receptor interaction and MTF-1 regulate the expression of ZnT2 (Slc30a2) in pancreatic acinar cells. Proc Natl Acad Sci USA107: 2818-2823, 2010. PMID: 20133611
- Haanes KA, Kowal JM, Arpino G, Lange SC, Moriyama Y, Pedersen PA, and Novak I. Role of vesicular nucleotide transporter VNUT (SLC17A9) in release of ATP from AR42J cells and mouse pancreatic acinar cells. Purinergic Signal 10: 431-440, 2014. PMID: 24488439
- Haanes KA and Novak I. ATP storage and uptake by isolated pancreatic zymogen granules. Biochem J 429: 303-311, 2010. PMID: 20441565
- Hamann M, Gibson A, Davies N, Jowett A, Walhin JP, Partington L, Affleck K, Trezise D, and Main M. Human ClCa1 modulates anionic conduction of calcium-dependent chloride currents. J Physiol 587: 2255-2274, 2009. PMID: 19307298
- Hirano Y, Hisatomi A, Ohara K, and Noguchi H. The effects of FK506 and cyclosporine on the exocrine function of the rat pancreas. Transplantation 54: 883-887, 1992. PMID: 1279850
- Huang L and Tepaamorndech S. The SLC30 family of zinc transporters - a review of current understanding of their biological and pathophysiological roles. Molecular aspects of medicine 34: 548-560, 2013. PMID: 23506888
- Huang P, Zou Y, Zhong XZ, Cao Q, Zhao K, Zhu MX, Murrell-Lagnado R, and Dong XP. P2X4 forms functional ATP-activated cation channels on lysosomal membranes regulated by luminal pH. J Biol Chem 289: 17658-17667, 2014. PMID: 23506888
- Ikeda M, Beitz E, Kozono D, Guggino WB, Agre P, and Yasui M. Characterization of aquaporin-6 as a nitrate channel in mammalian cells. Requirement of pore-lining residue threonine 63. J Biol Chem 277: 39873-39879, 2002. PMID: 12177001
- Jentsch TJ, Maritzen T, Keating DJ, Zdebik AA, and Thévenod F. ClC-3--a granular anion transporter involved in insulin secretion? Cell Metab 12: 307-308; author reply 309-310, 2010. PMID: 20889118
- Jespersen T, Grunnet M, and Olesen SP. The KCNQ1 potassium channel: from gene to physiological function. Physiology (Bethesda) 20: 408-416, 2005. PMID: 16287990
- Johnson RG, Jr. Accumulation of biological amines into chromaffin granules: a model for hormone and neurotransmitter transport. Physiol Rev 68: 232-307, 1988. PMID: 2892215
- Johnson RG, Jr. Proton pumps and chemiosmotic coupling as a generalized mechanism for neurotransmitter and hormone transport. Ann N Y Acad Sci 493: 162-177, 1987. PMID: 2884918
- Jou MY, Hall AG, Philipps AF, Kelleher SL, and Lonnerdal B. Tissue-specific alterations in zinc transporter expression in intestine and liver reflect a threshold for homeostatic compensation during dietary zinc deficiency in weanling rats. J Nutrition 139: 835-841, 2009. PMID: 19297427
- Kataoka M, Fukura Y, Shinohara Y, and Baba Y. Analysis of mitochondrial membrane potential in the cells by microchip flow cytometry. Electrophoresis 26: 3025-3031, 2005. PMID: 16078196
- Kelly ML, Abu-Hamdah R, Jeremic A, Cho SJ, Ilie AE, and Jena BP. Patch clamped single pancreatic zymogen granules: direct measurements of ion channel activities at the granule membrane. Pancreatology 5: 443-449, 2005. PMID: 15985770
- Kunzelmann K, Tian Y, Martins JR, Faria D, Kongsuphol P, Ousingsawat J, Thevenod F, Roussa E, Rock J, and Schreiber R. Anoctamins. Pflugers Arch 462: 195-208, 2011. PMID: 21607626
- Larsen KS and Auld DS. Characterization of an inhibitory metal binding site in carboxypeptidase A. Biochemistry 30: 2613-2618, 1991. PMID: 2001351
- Lee JE, Park HS, Uhm DY, and Kim SJ. Effects of KCNQ1 channel blocker, 293B, on the acetylcholine-induced Cl- secretion of rat pancreatic acini. Pancreas 28: 435-442, 2004. PMID: 15097862
- Lee WK, Bork U, Gholamrezaei F, and Thévenod F. Cd2+-induced cytochrome c release in apoptotic proximal tubule cells: role of mitochondrial permeability transition pore and Ca2+ uniporter. Am J Physiol Renal Physiol 288: F27-F39, 2005. PMID: 15339793
- Lee WK, Braun M, Langeluddecke C, and Thévenod F. Cyclosporin a, but not FK506, induces osmotic lysis of pancreas zymogen granules, intra-acinar enzyme release, and lysosome instability by activating K+ channel. Pancreas 41: 596-604, 2012. PMID: 22158076
- Lee WK, Torchalski B, Roussa E, and Thévenod F. Evidence for KCNQ1 K+ channel expression in rat zymogen granule membranes and involvement in cholecystokinin-induced pancreatic acinar secretion. Am J Physiol Cell Physiol 294: C879-C892, 2008. PMID: 18216164
- Lemaire K, Chimienti F, and Schuit F. Zinc transporters and their role in the pancreatic beta-cell. J Diabetes Invest 3: 202-211, 2012. PMID: 24843567
- Lerch MM and Gorelick FS. Models of acute and chronic pancreatitis. Gastroenterology 144: 1180-1193, 2013. PMID: 23622127
- Lerche C, Bruhova I, Lerche H, Steinmeyer K, Wei AD, Strutz-Seebohm N, Lang F, Busch AE, Zhorov BS, and Seebohm G. Chromanol 293B binding in KCNQ1 (Kv7.1) channels involves electrostatic interactions with a potassium ion in the selectivity filter. Mol Pharmacol 71: 1503-1511, 2007. PMID: 17347319
- Lesage F and Lazdunski M. Molecular and functional properties of two-pore-domain potassium channels. Am J Physiol Renal Physiol 279: F793-801, 2000. PMID: 11053038
- Liu K, Kozono D, Kato Y, Agre P, Hazama A, and Yasui M. Conversion of aquaporin 6 from an anion channel to a water-selective channel by a single amino acid substitution. Proc Natl Acad Sci USA 102: 2192-2197, 2005. PMID: 15671159
- Liuzzi JP, Bobo JA, Lichten LA, Samuelson DA, and Cousins RJ. Responsive transporter genes within the murine intestinal-pancreatic axis form a basis of zinc homeostasis. Proc Natl Acad Sci USA 101: 14355-14360, 2004. PMID: 15381762
- Luo X, Zheng W, Yan M, Lee MG, and Muallem S. Multiple functional P2X and P2Y receptors in the luminal and basolateral membranes of pancreatic duct cells. Am J Physiol Cell Physiol 277: C205-215, 1999. PMID: 10444396
- Martin L, Lodemann U, Bondzio A, Gefeller EM, Vahjen W, Aschenbach JR, Zentek J, and Pieper R. A high amount of dietary zinc changes the expression of zinc transporters and metallothionein in jejunal epithelial cells in vitro and in vivo but does not prevent zinc accumulation in jejunal tissue of piglets. J Nutrition 143: 1205-1210, 2013. PMID: 23761649
- Matsuki-Fukushima M, Fujita-Yoshigaki J, Murakami M, Katsumata-Kato O, Yokoyama M, and Sugiya H. Involvement of AQP6 in the Mercury-sensitive osmotic lysis of rat parotid secretory granules. J Membr Biol 246: 209-214, 2013. PMID: 23183829
- Matsuki-Fukushima M, Hashimoto S, Shimono M, Satoh K, Fujita-Yoshigaki J, and Sugiya H. Presence and localization of aquaporin-6 in rat parotid acinar cells. Cell Tissue Res 332: 73-80, 2008. PMID: 18188600
- Matsuki M, Hashimoto S, Shimono M, Murakami M, Fujita-Yoshigaki J, Furuyama S, and Sugiya H. Involvement of aquaporin-5 water channel in osmoregulation in parotid secretory granules. J Membr Biol 203: 119-126, 2005. PMID: 15986091
- Matsuzaki T, Ablimit A, Suzuki T, Aoki T, Hagiwara H, and Takata K. Changes of aquaporin 5-distribution during release and reaccumulation of secretory granules in isoproterenol-treated mouse parotid gland. J Electron Microscopy 55: 183-189, 2006. PMID: 16899467
- Medhurst AD, Rennie G, Chapman CG, Meadows H, Duckworth MD, Kelsell RE, Gloger, II, and Pangalos MN. Distribution analysis of human two pore domain potassium channels in tissues of the central nervous system and periphery. Brain Res Mol Brain Res 86: 101-114, 2001. PMID: 11165377
- Meldolesi J. Membranes of pancreatic zymogen granules. Methods Enzymol 98: 67-75, 1983. PMID: 6669071
- Messenger SW, Falkowski MA, and Groblewski GE. Ca2+-regulated secretory granule exocytosis in pancreatic and parotid acinar cells. Cell Calcium 55: 369-375, 2014. PMID: 24742357
- Miyaji T, Sawada K, Omote H, and Moriyama Y. Divalent cation transport by vesicular nucleotide transporter. J Biol Chem 286: 42881-42887, 2011. PMID: 22052906
- Nezu A, Tanimura A, Morita T, Irie K, Yajima T, and Tojyo Y. Evidence that zymogen granules do not function as an intracellular Ca2+ store for the generation of the Ca2+ signal in rat parotid acinar cells. Biochem J 363: 59-66, 2002. PMID: 11903047
- Nguyen T, Chin WC, and Verdugo P. Role of Ca2+/K+ ion exchange in intracellular storage and release of Ca2+. Nature 395: 908-912, 1998. PMID: 9804425
- Niederau C, Van Dyke RW, Scharschmidt BF, and Grendell JH. Rat pancreatic zymogen granules. An actively acidified compartment. Gastroenterology 91: 1433-1442, 1986. PMID: 3021556
- Norlund R, Roos N, and Taljedal IB. Quantitative energy dispersive X-ray microanalysis of eight elements in pancreatic endocrine and exocrine cells after cryo-fixation. Bioscience Reports 7: 859-869, 1987. PMID: 3329534
- Novak I. ATP as a signaling molecule: the exocrine focus. News Physiol Sci 18: 12-17, 2003. PMID: 12531926
- Ohana E, Hoch E, Keasar C, Kambe T, Yifrach O, Hershfinkel M, and Sekler I. Identification of the Zn2+ binding site and mode of operation of a mammalian Zn2+ transporter. J Biol Chem 284: 17677-17686, 2009. PMID: 19366695
- Ohta E, Itoh T, Nemoto T, Kumagai J, Ko SB, Ishibashi K, Ohno M, Uchida K, Ohta A, Sohara E, Uchida S, Sasaki S, and Rai T. Pancreas-specific aquaporin 12 null mice showed increased susceptibility to caerulein-induced acute pancreatitis. Am J Physiol Cell Physiol 297: C1368-1378, 2009. PMID: 19726746
- Orci L, Ravazzola M, and Anderson RG. The condensing vacuole of exocrine cells is more acidic than the mature secretory vesicle. Nature 326: 77-79, 1987. PMID: 2434862
- Orlowski J and Grinstein S. Emerging roles of alkali cation/proton exchangers in organellar homeostasis. Curr Opin Cell Biol 19: 483-492, 2007. PMID: 17646094
- Palmiter RD, Cole TB, and Findley SD. ZnT-2, a mammalian protein that confers resistance to zinc by facilitating vesicular sequestration. EMBO J 15: 1784-1791, 1996. PMID: 8617223
- Paroutis P, Touret N, and Grinstein S. The pH of the secretory pathway: measurement, determinants, and regulation. Physiology (Bethesda) 19: 207-215, 2004.
- Patel AC, Brett TJ, and Holtzman MJ. The role of CLCA proteins in inflammatory airway disease. Annu Rev Physiol 71: 425-449, 2009. PMID: 18954282
- Pekas JC. Pancreatic incorporation of 65Zn and histidine-14C into secreted proteins of the pig. Am J Physiol 220: 799-803, 1971. PMID: 5545692
- Petersen OH, Gerasimenko OV, Tepikin AV, and Gerasimenko JV. Aberrant Ca2+ signalling through acidic calcium stores in pancreatic acinar cells. Cell Calcium 50: 193-199, 2011. PMID: 21435718
- Petersen OH, Tepikin AV, Gerasimenko JV, Gerasimenko OV, Sutton R, and Criddle DN. Fatty acids, alcohol and fatty acid ethyl esters: toxic Ca2+ signal generation and pancreatitis. Cell Calcium 45: 634-642, 2009. PMID: 19327825
- Poea-Guyon S, Ammar MR, Erard M, Amar M, Moreau AW, Fossier P, Gleize V, Vitale N, and Morel N. The V-ATPase membrane domain is a sensor of granular pH that controls the exocytotic machinery. J Cell Biol 203: 283-298, 2013. PMID: 24165939
- Rindler MJ, Xu CF, Gumper I, Smith NN, and Neubert TA. Proteomic analysis of pancreatic zymogen granules: identification of new granule proteins. J Proteome Res 6: 2978-2992, 2007. PMID: 17583932
- Roomans GM and Wei X. X-ray microanalysis of resting and stimulated rat pancreas. Acta Physiol Scand 124: 353-359, 1985. PMID: 4050472
- Roos N. A possible site of calcium regulation in rat exocrine pancreas cells: an X-ray microanalytical study. Scanning Microscopy 2: 323-329, 1988. PMID: 3368762
- Roussa E, Alper SL, and Thévenod F. Immunolocalization of anion exchanger AE2, Na+/H+ exchangers NHE1 and NHE4, and vacuolar type H+-ATPase in rat pancreas. J Histochem Cytochem 49: 463-474, 2001. PMID: 11259449
- Roussa E, Wittschen P, Wolff NA, Torchalski B, Gruber AD, and Thévenod F. Cellular distribution and subcellular localization of mCLCA1/2 in murine gastrointestinal epithelia. J Histochem Cytochem 58: 653-668, 2010. PMID: 20385786
- Sawada K, Echigo N, Juge N, Miyaji T, Otsuka M, Omote H, Yamamoto A, and Moriyama Y. Identification of a vesicular nucleotide transporter. Proc Natl Acad Sci USA 105: 5683-5686, 2008. PMID: 18375752
- Seve M, Chimienti F, Devergnas S, and Favier A. In silico identification and expression of SLC30 family genes: an expressed sequence tag data mining strategy for the characterization of zinc transporters' tissue expression. BMC Genomics 5: 32, 2004. PMID: 15154973
- Shusterman E, Beharier O, Shiri L, Zarivach R, Etzion Y, Campbell CR, Lee IH, Okabayashi K, Dinudom A, Cook DI, Katz A, and Moran A. ZnT-1 extrudes zinc from mammalian cells functioning as a Zn2+/H+ exchanger. Metallomics : integrated biometal science 6: 1656-1663, 2014. PMID: 24951051
- Sondergaard LG, Stoltenberg M, Doering P, Flyvbjerg A, and Rungby J. Zinc ions in the endocrine and exocrine pancreas of zinc deficient rats. Histology and Histopathology 21: 619-625, 2006. PMID: 16528672
- Stauber T and Jentsch TJ. Chloride in vesicular trafficking and function. Annu Rev Physiol 75: 453-477, 2013. PMID: 23092411
- Stobrawa SM, Breiderhoff T, Takamori S, Engel D, Schweizer M, Zdebik AA, Bosl MR, Ruether K, Jahn H, Draguhn A, Jahn R, and Jentsch TJ. Disruption of ClC-3, a chloride channel expressed on synaptic vesicles, leads to a loss of the hippocampus. Neuron 29: 185-196, 2001.
- Thévenod F. Ion channels in secretory granules of the pancreas and their role in exocytosis and release of secretory proteins. Am J Physiol Cell Physiol 283: C651-C672, 2002. PMID: 11182090
- Thévenod F, Chathadi KV, Jiang B, and Hopfer U. ATP-sensitive K+ conductance in pancreatic zymogen granules: block by glyburide and activation by diazoxide. J Membr Biol 129: 253-266, 1992. PMID: 1433277
- Thévenod F, Dehlinger-Kremer M, Kemmer TP, Christian AL, Potter BV, and Schulz I. Characterization of inositol 1,4,5-trisphosphate-sensitive (IsCaP) and -insensitive (IisCaP) nonmitochondrial Ca2+ pools in rat pancreatic acinar cells. J Membr Biol 109: 173-186, 1989. PMID: 2527996
- Thévenod F, Haase W, and Hopfer U. Large-scale purification of calf pancreatic zymogen granule membranes. Anal Biochem 202: 54-60, 1992. PMID: 1377890
- Thévenod F, Kemmer TP, Christian AL, and Schulz I. Characterization of MgATP-driven H+ uptake into a microsomal vesicle fraction from rat pancreatic acinar cells. J Membr Biol 107: 263-275, 1989. PMID: 2469802
- Thévenod F, Roussa E, Benos DJ, and Fuller CM. Relationship between a HCO3- -permeable conductance and a CLCA protein from rat pancreatic zymogen granules. Biochem Biophys Res Commun 300: 546-554, 2003. PMID: 12504118
- Thévenod F and Schulz I. H+-dependent calcium uptake into an IP3-sensitive calcium pool from rat parotid gland. Am J Physiol 255: G429-440, 1988. PMID: 3263053
- Thiemann A, Grunder S, Pusch M, and Jentsch TJ. A chloride channel widely expressed in epithelial and non-epithelial cells. Nature 356: 57-60, 1992. PMID: 1311421
- Thomas GP, Gerlach U, and Antzelevitch C. HMR 1556, a potent and selective blocker of slowly activating delayed rectifier potassium current. J Cardiovasc Pharmacol 41: 140-147, 2003. PMID: 12500032
- Thorn P, Lawrie AM, Smith PM, Gallacher DV, and Petersen OH. Ca2+ oscillations in pancreatic acinar cells: spatiotemporal relationships and functional implications. Cell Calcium 14: 746-757, 1993. PMID: 8131191
- Titievsky AV, Takeo T, Tepikin AV, and Petersen OH. Decrease of acidity inside zymogen granules inhibits acetylcholine- or inositol trisphosphate-evoked cytosolic Ca2+ spiking in pancreatic acinar cells. Pflugers Arch 432: 938-940, 1996. PMID: 8772146
- Vallee BL and Neurath H. Carboxypeptidase, a zinc metalloenzyme. J Biol Chem 217: 253-261, 1955. PMID: 13271390
- van de Put FH and Elliott AC. The endoplasmic reticulum can act as a functional Ca2+ store in all subcellular regions of the pancreatic acinar cell. J Biol Chem 272: 27764-27770, 1997. PMID: 9346920
- Vandecaetsbeek I, Vangheluwe P, Raeymaekers L, Wuytack F, and Vanoevelen J. The Ca2+ pumps of the endoplasmic reticulum and Golgi apparatus. Cold Spring Harbor Perspectives in Biology 3, 2011. PMID: 21441596
- Wang D, Epstein D, Khalaf O, Srinivasan S, Williamson WR, Fayyazuddin A, Quiocho FA, and Hiesinger PR. Ca2+-Calmodulin regulates SNARE assembly and spontaneous neurotransmitter release via v-ATPase subunit V0a1. J Cell Biol 205: 21-31, 2014. PMID: 24733584
- Williams JA. The nobel pancreas: a historical perspective. Gastroenterology 144: 1166-1169, 2013. PMID: 23622125
- Yang K, Ding YX, and Chin WC. K+-induced ion-exchanges trigger trypsin activation in pancreas acinar zymogen granules. Arch Biochem Biophys 459: 256-263, 2007. PMID: 17270141
- Yang YD, Cho H, Koo JY, Tak MH, Cho Y, Shim WS, Park SP, Lee J, Lee B, Kim BM, Raouf R, Shin YK, and Oh U. TMEM16A confers receptor-activated calcium-dependent chloride conductance. Nature 455: 1210-1215, 2008. PMID: 18724360
- Yasui M, Hazama A, Kwon TH, Nielsen S, Guggino WB, and Agre P. Rapid gating and anion permeability of an intracellular aquaporin. Nature 402: 184-187, 1999. PMID: 10647010
- Yoo SH. Role of secretory granules in inositol 1,4,5-trisphosphate-dependent Ca2+ signaling: from phytoplankton to mammals. Cell Calcium 50: 175-183, 2011. PMID: 21176957
- Yule DI. Cell Calcium. Special issue: Ca2+ signaling and secretory function. Preface. Cell Calcium 55: 281, 2014. PMID: 24813115
- Yule DI, Ernst SA, Ohnishi H, and Wojcikiewicz RJ. Evidence that zymogen granules are not a physiologically relevant calcium pool. Defining the distribution of inositol 1,4,5-trisphosphate receptors in pancreatic acinar cells. J Biol Chem 272: 9093-9098, 1997. PMID: 9083036
- Zhao H and Muallem S. Na+, K+, and Cl- transport in resting pancreatic acinar cells. J Gen Physiol 106: 1225-1242, 1995. PMID: 8786358
- Zheng D, Feeney GP, Handy RD, Hogstrand C, and Kille P. Uptake epithelia behave in a cell-centric and not systems homeostatic manner in response to zinc depletion and supplementation. Metallomics 6: 154-165, 2014. PMID: 24301558